PHYSIOLOGIE DE L'EXERCICE *
Les fibres musculaires *
Classification des fibres motrices *
La fibre lente (type I) *
La fibre intermédiaire (type IIa) *
La fibre rapide (type IIB) *
Physiologie des fibres motrices *
Transmission de l’excitation au muscle *
Les fuseaux neuro-musculaire et le système gamma *
Physiologie de l'exercice *
Le métabolisme de base *
L'effet dynamique spécifique *
COUT ENERGETIQUE DES ACTIVITES *
Dépense énergétique approximative des activités manuelles les plus courantes. *
Limites d'ordre quantitatif pour le transport d'une charge. *
Les substrats énergétiques *
1°) La glycolyse anaérobie alactique: ATP et CP *
2°) La glycolyse anaérobie lactique: le glycogène et l'acide lactique *
Le glycogène *
La surcharge glycogénique *
L'acide lactique *
Hypothèses de la diminution d’acide lactique dans un exercice sous-maximal *
3°) La glycolyse aérobie: les acides gras libres *
4°) la filière protéique: la néoglucogenèse *
Le métabolisme du glucose *
Fraction moyenne d'utilisation des différents substrats pendant l'exercice *
Substrats et rendements énergétiques maximum des différentes sources énergétiques dans la cellule musculaire *
Métabolisme oxydatif *
Les mécanismes d'adaptation à l'effort *
I/ adaptation cardiaque *
Le volume d'éjection systolique *
Le débit cardiaque *
La fréquence cardiaque *
Physiologie de l'adaptation de Fc à l'effort *
Le débit sanguin à l'effort du myocarde *
II/ Adaptation de la distribution sanguine régionale *
III/ Adaptation de la distribution sanguine locale *
L'augmentation du retour veineux *
IV/ Adaptation orthosympathique et endocrinienne *
V/ Adaptation de la tension artérielle *
Conditions tensionnelles d'arrêt de l'exercice *
Etudes réalisées par Astrand sur l'évolution de la tension suivant le type d'effort *
Modifications tensionnelles lors de la récupération *
VI/ Adaptation ventilatoire *
Le second souffle *
La production de dioxyde de carbone *
Le quotient respiratoire *
QR = VCO2 / VO2 *
Gaz du sang et lactate *
Discussion à propos du seuil anaérobie *
Evolution des paramètres respiratoires *
La ventilation pulmonaire *
VII/ Adaptation de la consommation d'oxygène *
La dette d'oxygène *
VIII/ Amélioration de la différence artério-veineuse *
IX/ Adaptation de la courbe de dissociation de l'oxyhémoglobine (HbO2) *
L'hémoglobine *
La myoglobine *
X/ Les facteurs limitants de la performance *
XI/ La fatigue *
La thermorégulation *
La chaleur et les pertes d'eau pendant l'exercice *
Les diurétiques *
Les électrolytes *
Prévention des troubles liés à la chaleur *
La boisson de l'effort *
Concentration de glucose selon la température ambiante: *
Le refroidissement excessif *
Les symptômes du coup de chaleur *
Les soins au sujet victime d'un coup de chaleur *
L'acclimatation *
Le sauna *
La phase de réchauffement *
Effets de la phase de réchauffement *
La phase de refroidissement *
Effets de la phase de refroidissement *
Effets du sauna *
Contre-indications *
Intérêt du sauna pour le sportif *
L'adaptation de l'organisme au froid *
La récupération après l'effort *
Paiement de la dette d'oxygène *
Restauration de la myoglobine *
Récupération de l'ATP *
Restauration de la phosphocréatine *
Rétablissement des réserves de phosphagènes du muscle: 2 à 5 mn *
Restauration du glycogène *
Exercices épuisants de longue durée *
Exercices épuisants de courte durée *
Elimination de l'acide lactique *
Le catabolisme de l'AL *
La diététique de l'effort *
Le repas pré-compétition *
L'alimentation pendant la compétition *
La boisson de l'effort *
Le repas post-compétition *
Les troubles de l'effort *
Lésions musculaires *
Causes de la douleur musculaire et de la raideur *
Les courbatures *
La crampe *
L’élongation *
La déchirure musculaire *
Les malaises *
Le malaise hypoglycémique *
Le malaise vagal *
Troubles cardiaques pendant l'exercice *
Insuffisance cardiaque aiguë des cœurs sains *
Troubles du fonctionnement du tissu nodal *
Les problèmes de !a mort subite du sportif après autopsie blanche (négative) *
Le pseudo-collapsus cardio-périphérique du sportif *
Physiopathologie de la mort subite du sportif *
Les manœuvres vagales à utiliser en cas de tachycardie (flutter, WPW…) *
Attitude à adopter en cas de malaise cardiaque du patient *
Précautions à prendre durant la réanimation cardio-pulmonaire *
- Desserrer les vêtements et la ceinture de manière à libérer les voies aériennes *
Troubles du système épuratoire dus à l'exercice *
Hémoglobinurie d'effort *
Myoglobinurie d’effort et rhabdomyolyse *
Troubles du système tampon dus à l'exercice *
Acidose métabolique *
Alcalose métabolique *
Acidose respiratoire *
Alcalose respiratoire *
Fractures spontanées (ou de fatigue) du sportif *
La fonction physiologique de toute fibre
sensitive ou motrice se traduit par la variation du potentiel de repos. Ce
potentiel de repos est à -70 mV, la fibre étant polarisée positivement à
l'extérieur et négativement à l'intérieur.
La réalisation d'un influx moteur va suivre
la loi du "tout ou rien", c'est à dire que l'influx devra avoir une
variation minima de 100 mV pour dépolariser la fibre, le potentiel de fibre
passant à +30 mV. La vitesse de dépolarisation varie en fonction de la structure
de la fibre de 15 à
 La fibre revient ensuite à son état initial en passant par une période
d'hyperpolarisation réfractaire absolue de 1 ms pendant laquelle elle n'est plus
excitable et une période réfractaire relative pouvant durer jusqu'à 15 ms et
pendant laquelle la fibre n'est excitable que par un nouvel influx plus intense
que le précédent.
La fibre revient ensuite à son état initial en passant par une période
d'hyperpolarisation réfractaire absolue de 1 ms pendant laquelle elle n'est plus
excitable et une période réfractaire relative pouvant durer jusqu'à 15 ms et
pendant laquelle la fibre n'est excitable que par un nouvel influx plus intense
que le précédent.
En tenant compte da la période réfractaire,
on calcule qu'un nerf ne peut pas transmettre des influx supérieurs à 1000 Hz
Les fibres sont classées en plusieurs types
principaux:
F aa fibres nerveuses motrices
F fibres Ia proprio-sensitives des fuseaux musculaires, 80 à 120 m/s
F fibres Ib proprio-sensitives des organes de Golgi tendineux
F ab fibres sensitives de grand diamètre, myélinisées, très rapides,
afférentes des récepteurs de la peau, inhibitrices, antalgiques, 60 m/s
F ag fibres motrices efférentes des fuseaux musculaires
F ad fibres sensitives de petit diamètre, myélinisées, moins rapides que ab ,
nociceptives, provoquant une douleur rapide, fulgurante, à type de piqûre,
afférentes des récepteurs sensitifs de la peau pour la chaleur, le froid, la
douleur, 15 à 25 m/s
F B fibres végétatives préganglionnaires, 3 à 15 m/s
F C fibres sensitives de petit diamètre, non myélinisées, lentes,
nociceptives provoquant une douleur tenace à type de brûlure
F différents types d'autres fibres (postganglionnaires, fibres afférentes du
nerf grand sympathique, fibres sensitives de la peau, fibres des racines
postérieures)
Classification des fibres motrices
Elles sont classées en trois types
principaux dont le métabolisme est différent :
F rapides
F intermédiaires
F lentes

La cellule musculaire est en fait une fibre
constituée de plusieurs noyaux situés en périphérie et de plusieurs milliers de
myofibrilles parallèles, entourées de mitochondries et situées dans le
sarcoplasme avec d'autres constituants cellulaires.
Les fibres lentes sont majoritaires dans les
muscles de posture (muscles du rachis, abdominaux), les fibres rapides sont
majoritaires dans les muscles travaillant essentiellement de manière dynamique
(quadriceps, muscles des yeux, des doigts etc…). De la même manière, les hommes
et femmes sédentaires possèdent 45 à 55% de fibres lentes alors qu'un sportif
d'endurance possédera une musculature majoritairement en fibres lentes et qu'un
sprinter possédera une musculature majoritairement en fibres rapides.
L'entraînement permet de plus ou moins spécialiser un type de fibres. Chez les
sportifs entraînés "en force", la surface de section des différentes
fibres peut surpasser de 45% celle des athlètes d’endurance ou d’individus
sédentaires de même âge, car l’entraînement en force et en puissance stimule le
développement de l’appareil contractile. Les athlètes mâles ont habituellement
des fibres plus volumineuses que les athlètes femelles. Cependant,
l’identification d’un nombre prédominant de fibres n’a pas d’incidence sur la
performance, car c’est l’interaction optimale de tous les systèmes qui est
importante.
Les motoneurones de la corne antérieure sont
du type A. Lorsque leur diamètre varie entre l0 et 20 µm, ce sont des fibres Aa . On
distingue aussi d’autres fibres de type A, les fibres efférentes gamma (g ), dont le
diamètre est inférieur à 10 µm et la vitesse de conduction moitié moindre. Ces
fibres innervent des capteurs intramusculaires sensibles à l’étirement, dont le
rôle est de détecter les minimes variations de longueur des fibres musculaires.
F Les fibres motrices rapides sont sollicitées par la stimulation
électrique normalement juste avant ou en même temps que les fibres sensitives ab .
F Plus le diamètre d'une fibre est grand, plus vite elle est dépolarisée.
F En pratique les fibres sensitives sont excitées par des influx pouvant
aller jusqu'à 300 Hz de fréquence et les fibres musculaires par des influx de
100 Hz de fréquence
maximum.

schéma
des courbes de Howson
 l fibre rouge de contraction
lente
l fibre rouge de contraction
lente
l
très résistante à la fatigue
l
activité tonique
l
métabolisme aérobique oxydatif
l
mitochondries
l
les fibres I sont entourées d'un réseau capillaire très dense pouvant atteindre
La fibre intermédiaire (type IIa)
 l fibre intermédiaire de
contraction rapide
l fibre intermédiaire de
contraction rapide
l
résistante à l’activité intermédiaire
l
tonico-phasique
La fibre de type IIa est dite intermédiaire
du fait qu’elle est relativement rapide et qu’elle présente une bonne capacité
aérobie (haut niveau d’activité de l’enzyme S.D.H.) et anaérobie (haut niveau
l’activité de l’enzyme P.F.K.) alors que les fibres du type IIb ont le plus
grand potentiel anaérobie ; ce sont les fibres FG (fast glycolitic). La fibre
du type IIc est rare et non différenciée : c’est probablement une fibre en voie
de réinnervation ou de transformation.
 l fibre blanche de
contraction rapide typiquement phasique
l fibre blanche de
contraction rapide typiquement phasique
l
métabolisme anaérobique pauvre en mitochondries
l
moins oxydative et plus anaérobique
l
développe une grande puissance mais de courte durée
l
très fatigable.
l
riche en glycogène
L'entraînement permet aux fibres rapides
d'améliorer leur potentiel métabolique aérobie sans entrer en concurrence avec les
fibres lentes et de déterminer ainsi une augmentation de la consommation d'O2
Physiologie des fibres motrices
 Transmission de l’excitation au muscle
Transmission de l’excitation au muscle
L’influx part de la cellule ganglionnaire
motrice jusqu'aux plaques motrices, chaque
plaque motrice commandant plusieurs fibres.
La plaque motrice excitée libère les molécules d'acethylcholine qui vont
permettre la contraction des fibres.
Notons que plus le mouvement est précis,
plus il y a d'axones innervant le muscle et moins il y a de fibres par plaque
motrice. Un muscle de l'œil alignera 10 fibres maximum par plaque, alors qu'un
muscle squelettique alignera jusqu'à 2000 fibres par plaque.
Notons que le muscle lisse nécessite un
temps de contraction plus grand que le muscle strié.
L'augmentation du nombre de fibres
musculaires recrutées dans une contraction permet de diminuer la dépense
énergétique et musculaire. La corticalisation des acquis effectuée, permet de
faire passer ce contrôle au second degré et de libérer la concentration et la
réflexion pour d'autres tâches.
 L'hyperplasie des fibres musculaires n'est pas prouvée chez l'humain.
L'hyperplasie des fibres musculaires n'est pas prouvée chez l'humain.
Chaque fibre musculaire striée présente
généralement une seule jonction neuro-musculaire, concernant le même type de
fibres. Toutes les fibres musculaires d’une unité motrice sont du même type
mais ces fibres musculaires ne sont pas groupées mais entremêlées avec d’autres
fibres de type différents et dépendant d’autres unités motrices. Les fibres
rapides sont innervées par de gros motoneurones, les fibres lentes par des
petits neurones. La stimulation d'un seul motoneurone alpha peut faire
contracter jusqu’à 300 fibres musculaires.
Les terminaisons d’un nerf innervent au
moins une des 250 millions de fibres musculaires de l’organisme. Comme il n'y a
qu’environ 420 000 fibres nerveuses motrices, chacune d'entre elles innerve
donc plusieurs fibres musculaires.
Les neurones conduisent l’influx dans une
seule direction, dite centrifuge. La vitesse de conduction d’une fibre nerveuse
est proportionnelle au diamètre de la fibre et à l’épaisseur de sa gaine.
 La force d’une contraction varie par nombre d’unités motrices recrutées
et par augmentation des fréquences. Pour des puissances croissantes, les fibres
musculaires à contraction rapide sont progressivement sollicitées. Elles ont
une aptitude élevée pour le métabolisme anaérobie et consomment le glycogène de
façon accélérée. Leur stock se réduit douze à treize fois plus rapidement que
celui des fibres à contraction lente, car la contraction dynamique peut
multiplier par 100 le métabolisme du muscle. Cependant après 9 à 12 heures
d'exercice, le seul glycogène restant est celui des fibres rapides inutilisées.
La répétition prolongée de contractions musculaires entraîne une perte d'ions
potassium et de calcium dans la cellule musculaire. La séparation des ponts
d'actine-myosine nécessitant de l'ATP, qui n'est plus fourni pas manque de
glycogène, ne se produit pas et induit une contracture; c'est pourquoi, la
contracture intéresse de manière privilégiée les fibres rapides.
La force d’une contraction varie par nombre d’unités motrices recrutées
et par augmentation des fréquences. Pour des puissances croissantes, les fibres
musculaires à contraction rapide sont progressivement sollicitées. Elles ont
une aptitude élevée pour le métabolisme anaérobie et consomment le glycogène de
façon accélérée. Leur stock se réduit douze à treize fois plus rapidement que
celui des fibres à contraction lente, car la contraction dynamique peut
multiplier par 100 le métabolisme du muscle. Cependant après 9 à 12 heures
d'exercice, le seul glycogène restant est celui des fibres rapides inutilisées.
La répétition prolongée de contractions musculaires entraîne une perte d'ions
potassium et de calcium dans la cellule musculaire. La séparation des ponts
d'actine-myosine nécessitant de l'ATP, qui n'est plus fourni pas manque de
glycogène, ne se produit pas et induit une contracture; c'est pourquoi, la
contracture intéresse de manière privilégiée les fibres rapides.
Noter que les fibres lentes ST ont une
grande capacité aérobie, mais également une petite capacité anaérobie.
L’inverse est vrai pour les fibres rapides FT.
Les
fuseaux neuro-musculaires et le système gamma
Les fuseaux neuro-musculaires transmettent
l’information afférente sur la longueur et la tension des fibres musculaires.
Leur fonction principale est de réagir à l’étirement du muscle en initiant, par
voie réflexe, une contraction plus forte qui réduit l’étirement. Le fuseau
neuro-musculaire est lié, en parallèle, aux fibres musculaires extrafusales. A
l’intérieur du fuseau, on distingue deux types de fibres musculaires spéciales
dites intrafusales:
 £ Les fibres à sac nucléaire
sont de bonne dimension et renferment plusieurs noyaux dans la région
équatoriale. On en compte habituellement deux par fuseau.
£ Les fibres à sac nucléaire
sont de bonne dimension et renferment plusieurs noyaux dans la région
équatoriale. On en compte habituellement deux par fuseau.
£ L’autre type de fibres intrafusales renferme
plusieurs noyaux, étalés tout le long de la fibre.
Les régions polaires des fibres intrafusales
sont striées (elles contiennent de l’actine et de la myosine) et sont capables
de se contracter.
Ces fuseaux neuro-musculaires reçoivent
l’innervation des trois types de fibres nerveuses : deux sont afférentes ou
sensitives et une est efférente ou motrice.
 £ Une afférence primaire
s'enroule autour de la région équatoriale. C’est la fibre annulospiralée qui
réagit directement à l’étirement du muscle; la fréquence de ses potentiels
d’action est proportionnelle au degré d’étirement.
£ Une afférence primaire
s'enroule autour de la région équatoriale. C’est la fibre annulospiralée qui
réagit directement à l’étirement du muscle; la fréquence de ses potentiels
d’action est proportionnelle au degré d’étirement.
£ L’autre type d’afférence, qui est plus petite, et
dont les terminaisons s’étalent comme un bouquet, innerve principalement les
fibres à chaîne nucléaire, bien qu’on la trouve aussi au niveau des fibres à
sac nucléaire. Ces dernières afférences sont moins sensibles à l’étirement que
les premières.
L’activation des afférences annulospiralées
et en bouquet déclenche une volée d’influx qui emprunte les racines
postérieures (voie sensorielle) pour aller stimuler de façon réflexe les
motoneurones localisés dans la moelle épinière. Le muscle se contracte donc
plus fortement, se raccourcit et annule ainsi la stimulation des fuseaux
neuro-musculaires.
La mince fibre gamma efférente dont les
terminaisons innervent les régions polaires contracte des fibres musculaires
intrafusales. Ces fibres, dont l’activation relève des centres nerveux
supérieurs, complètent le mécanisme. La stimulation des fibres gamma fait
contracter les régions contractiles des fibres intrafusales, ajustant ainsi la
sensibilité du fuseau neuro-musculaire pour toute longueur musculaire.
Les organes neuro-tendineux (ou organes de
Golgi) sont reliés en série à près de 25 fibres musculaires extrafusales. On
retrouve aussi des récepteurs dans les ligaments des articulations; ils sont
d'abord et avant tout des détecteurs de la tension et non de la longueur
musculaire. La fonction première des organes neuro-tendineux est de protéger le
muscle et son tissu conjonctif, de blessures dues à une trop grande charge.
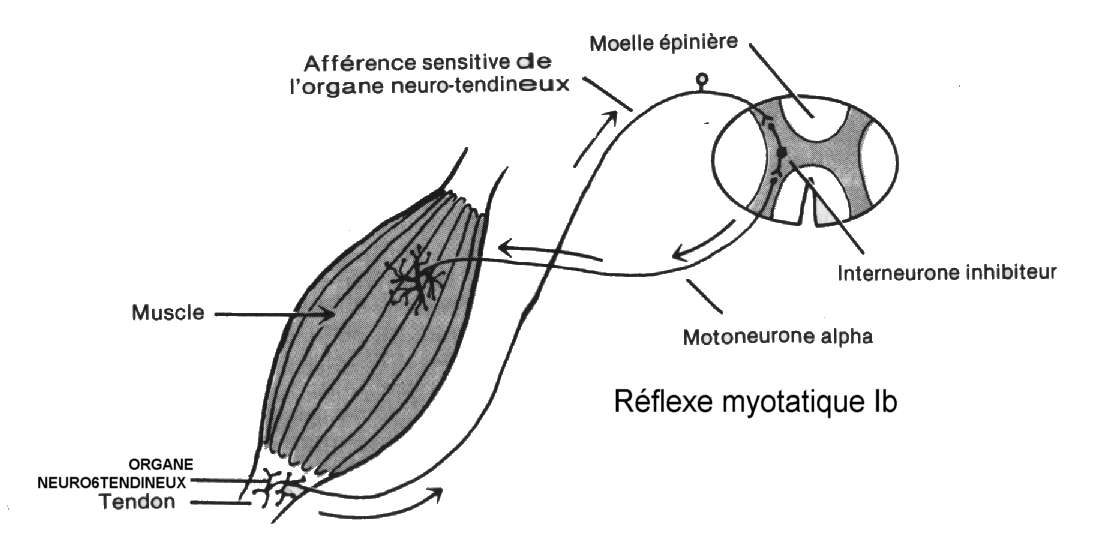
Ce sont les récepteurs propriocepteurs qui
transmettent rapidement l’information relative à la dynamique musculaire par
des fibres spécialisées. Les corpuscules de Pacini sont de petites structures
elliptiques localisées près des organes neuro-tendineux. Ils détectent les
variations de mouvements et de pression.
Le métabolisme de base est le besoin en
calories nécessité par l'organisme pour assurer son fonctionnement pendant une
journée Le métabolisme de base représente le niveau métabolique d'un individu
au repos, à jeun depuis 14 à18 heures. On considère que le MB est celui de
l'individu couché éveillé ou endormi.
 Le métabolisme de base s'adapte aux contraintes imposées et peut
diminuer de 50%. Les régimes hypocaloriques engendrent ainsi une baisse
considérable du métabolisme de base entraînant des prises de poids rebond et
l'échec du traitement. Un sujet équilibré à 2800 Kcal/jour soumis à un régime
de 1300 Kcal/jour verra son métabolisme chuter à 80% en 3 semaines et s'adapter
à ces 1300 Kcal/jour en 5 à 7 mois.
Le métabolisme de base s'adapte aux contraintes imposées et peut
diminuer de 50%. Les régimes hypocaloriques engendrent ainsi une baisse
considérable du métabolisme de base entraînant des prises de poids rebond et
l'échec du traitement. Un sujet équilibré à 2800 Kcal/jour soumis à un régime
de 1300 Kcal/jour verra son métabolisme chuter à 80% en 3 semaines et s'adapter
à ces 1300 Kcal/jour en 5 à 7 mois.
Le métabolisme de base s'exprime en Kj
(kilojoule) par cm2 de surface corporelle. [1 calorie = 4,18
joules].
métabolisme de repos en fonction de l'âge et du sexe
(Altman, PL & Dittmer, DS Metabolism, Bethseda, Md USA)
Il faut noter:
- Le métabolisme de base de la femme
apparaît de 5 à 10 % plus faible que celui de l'homme du fait de la plus grande
importance en graisse de sa masse corporelle. Par unité de masse maigre, le
métabolisme de base est identique.
- La dépense moyenne d'un individu
sédentaire âgé entre 23 et 50 ans varie de 2100 à 2700 Kcal (8790 à 11300 Kj).
Le métabolisme de l'homme s'élève à 1 Kcal par kg de poids corporel par heure.
- Les besoins énergétiques quotidiens d'un
sujet sédentaire d'âge moyen sont de 35 Kcal par kg de poids de corps.
- Le SNC consomme 20% du métabolisme basal
de l'homme pour 2 à 8% dans les autres espèces.
- Le métabolisme
de base est abaissé sous les tropiques.
- Le métabolisme
de base est plus élevé en hiver
- Le métabolisme
hormonal augmente le métabolisme de base
- Le métabolisme
de base du sportif est supérieur de 5% à celui du non sportif.
- L'enfant a un métabolisme plus élevé de 25
à 30% et nécessite 2.5 g/kg de poids de corps en protéines.
- Une heure de repos consomme en moyenne 100
Kcal.
- Quand un obèse perd 10% de son poids, ses
dépenses énergétiques baissent spontanément jusqu'à 16%.
F
Les dépenses de base assurent le fonctionnement des viscères et le maintien des
équilibres ioniques, elle coûtent une moyenne de 1400 Kcal.
F
Les dépenses de semi-repos assurent une activité musculaire modérée, la
thermorégulation, la digestion, elles coûtent une moyenne de 500 Kcal.
F
Les dépenses de travail léger coûtent 50 à 100 Kcal/heure
Les dépenses de travail moyen coûtent 100 à
300 Kcal/h
Les dépenses de travail lourd coûtent 300 à
450 Kcal/h
Les dépenses de travail intense coûtent 500
à 600 Kcal/h
 Il s'agit des dépenses énergétiques occasionnées par le mécanisme
digestif. De ce fait, plus il y a de repas dans la journée, plus les dépenses
énergétiques engendrées par la digestion sont importantes. Il est donc
préférable de répartir sa ration alimentaire sur plusieurs repas, de manière à
augmenter les dépenses énergétiques.
Il s'agit des dépenses énergétiques occasionnées par le mécanisme
digestif. De ce fait, plus il y a de repas dans la journée, plus les dépenses
énergétiques engendrées par la digestion sont importantes. Il est donc
préférable de répartir sa ration alimentaire sur plusieurs repas, de manière à
augmenter les dépenses énergétiques.
COUT ENERGETIQUE DES ACTIVITES
(EW Bannister & SR Brown The relative energy requirements of
physical activity)
Les dépenses sportives peuvent être
appréhendées par le tableau suivant:
Table sportive de
Kesterer et Knipping
|
Course à pied |
Kcal/h |
|
Vitesse |
500 |
|
Demi-fond |
930 |
|
Fond |
750 |
|
Marathon |
700 |
|
Cyclisme |
|
|
Sur piste |
220 |
|
Derrière entraîneur |
330 |
|
Sur route |
360 |
|
Sur route avec vent debout |
600 |
|
Ski |
|
|
Fond |
750 |
|
Alpin |
960 |
|
Tennis simple |
800 |
|
Tennis double |
350 |
|
Lancers |
460 |
|
Saut |
400 |
|
Aviron |
500 |
|
Lutte |
900 |
|
Boxe |
600 |
|
Haltérophilie |
450 |
|
Escrime |
600 |
|
Natation vitesse |
700 |
|
Natation fond |
450 |
|
Patinage artistique |
600 |
|
Patinage vitesse |
720 |
|
Basket-ball |
600 |
|
Foot-ball |
400 |
|
Water-polo |
600 |
|
Hand-ball |
500 |
|
Rugby |
500 |
Sport Femme Homme
Badminton 5.5
Kcal/mn 6.5
Cyclotourisme 5 5.9
Golf 4.4 5.2
Jogging
Marche plat lente 2.2 2.6
Marche
Idem +
Natation détente 7.7 9.1
Natation sportive 16 25
Squash 14.3 16
Tennis simple 9.4 11.1
Ces calculs sont variables et aléatoires, car
ils diffèrent par la tension nerveuse, l'entraînement, l'intensité etc... et
peuvent varier de 1 à 4.
Dépenses
énergétiques quotidiennes selon l'activité ou la profession
(I Katch, WD Mc ardle, Nutrition, Masse corporelle et
activité physique)
|
|
KJ.min /kg |
|
ascension de collines avec |
0.615 |
|
idem avec |
0.586 |
|
idem avec |
0.540 |
|
idem sans charge |
0.506 |
|
Badminton |
0.406 |
|
Balayage de tapis (F) |
0.188 |
|
Balayage de tapis (H) |
0.200 |
|
Basket ball |
0.578 |
|
Billard |
0.176 |
|
Boulangerie (F) |
0.146 |
|
Boxe, combat |
0.929 |
|
Boxe, entraînement |
0.578 |
|
Canotage, complet |
0.431 |
|
Canotage, loisir |
0.184 |
|
Cheval, course |
0.573 |
|
Cheval, entretien |
0.536 |
|
Cheval, marcher |
0.172 |
|
Cheval, trotter |
0.460 |
|
Circuit, entraîn |
0.774 |
|
Commis d'entrepôt |
0.226 |
|
Confection vêtements |
|
|
-Coudre machine |
0.188 |
|
-Coudre main |
0.134 |
|
-Couper |
0.172 |
|
-Presser |
0.260 |
|
Cordonnerie |
0.188 |
|
Cross country |
0.682 |
|
Course 3,18 min/Km |
1.210 |
|
Course3.45 min/km |
1.055 |
|
Course 4.20 min/km |
0.954 |
|
Course 5 min/km |
0.870 |
|
Course 5.30 min |
0.808 |
|
Course 7 min/Km |
0.565 |
|
Creuser tranchées |
0.607 |
|
Cricket, frappe |
0.347 |
|
Cricket, lancer |
0.377 |
|
Croquet |
0.247 |
|
Cuisson (F) |
0.188 |
|
Cuisson (H) |
0.201 |
|
Cyclisme, compet |
0.707 |
|
-loisir |
0.419 |
|
-loisir |
0.267 |
|
Dactylo, électrique |
0.113 |
|
Dactylo, manuelle |
0.130 |
|
|
Danse, chorégraphie, twist |
0.703 |
|
|
Danse, sociale |
0.213 |
|
|
Décapage |
0.264 |
|
|
Dessin debout |
0.150 |
|
|
Ecriture assis |
0.121 |
|
|
Extraction charbon |
0.368 |
|
|
Forage du sol |
0.452 |
|
|
Pelletage charbon |
0.552 |
|
|
Football américain |
0.456 |
|
|
Frottage plancher (F) |
0.452 |
|
|
Golf |
0.356 |
|
|
Gymnastique |
0.276 |
|
|
Hockey sur gazon |
0.561 |
|
|
Horlogerie |
0.105 |
|
|
Imprimerie |
0.146 |
|
|
Jardinage, border |
0.322 |
|
|
-creuser |
0.527 |
|
|
-rateler |
0.226 |
|
|
-tondre |
0.469 |
|
|
Jeu de cartes |
0.105 |
|
|
Judo |
0.816 |
|
|
Lavage plancher (F) |
0.260 |
|
|
Lavage plancher (H) |
0.243 |
|
|
Machines, percer |
0.272 |
|
|
-tourner |
0.200 |
|
|
-presser |
0.218 |
|
|
-plier métal |
0.368 |
|
|
-souder |
0.200 |
|
|
Manger assis |
0.218 |
|
|
Marche, allure régulière |
0.096 |
|
|
-champ labouré |
0.322 |
|
|
-champs et collines |
0.343 |
|
|
-chemin d'asphalte |
0.335 |
|
|
-sentier gazonné |
0.339 |
|
|
-militaire rapide |
0.594 |
|
|
marché (faire le) F |
0.260 |
|
|
idem H |
0.243 |
|
|
Menuiserie |
0.218 |
|
|
Métallurgie, conditionner |
0.373 |
|
|
-enlever machefer |
0.745 |
|
|
-forger |
0.419 |
|
|
-incliner moules- |
0.385 |
|
|
-laminer |
0.607 |
|
|
--rouler à la main |
0.573 |
|
|
-surveiller haut fourneau |
0.527 |
|
|
Musique, accordéon assis |
0.134 |
|
|
-batterie, assis |
0.276 |
|
|
-bois, assis |
0.134 |
|
|
-conduite orchestre |
0.163 |
|
|
-cor, assis |
0.121 |
|
|
-flûte, assis |
0.147 |
|
|
-orgue, assis |
0.222 |
|
|
-piano, assis |
0.167 |
|
|
-trompette, debout |
0.130 |
|
|
-violon, assis |
0.188 |
|
|
-violoncelle, assis |
0.172 |
|
|
Nage, brasse |
0.678 |
|
|
-crawl lent |
0.536 |
|
|
-crawl rapide |
0.653 |
|
|
-dos crawlé |
0.707 |
|
|
-marinière |
0.511 |
|
|
-sur place, normal |
0.260 |
|
|
-sur place, rapide |
0.711 |
|
|
Nettoyage (F) |
0.260 |
|
|
Nettoyage (H) |
0.243 |
|
|
Nettoyage fenêtres (F) |
0.247 |
|
|
idem (H) |
0.243 |
|
|
Papier peint, pose |
0.201 |
|
|
Pêche |
0.260 |
|
|
Peinture, extérieur |
0.322 |
|
|
Peinture, intérieur |
0.142 |
|
|
Pelleterie |
0.347 |
|
|
Planter jeunes plants |
0.293 |
|
|
Platrage |
0.326 |
|
|
Plongée sous-marine, modérée |
0.862 |
|
|
idem, très active |
1.155 |
|
|
Positions assise, tranquille |
0.088 |
|
|
Position debout, tranquille (F) |
0.105 |
|
|
Position debout, tranquille (H) |
0.113 |
|
|
Raquettes, marche sur neige poudreuse |
0.695 |
|
|
0.159 |
||
|
Repassage (F) |
0.138 |
|
|
Repassage (H) |
0.268 |
|
|
Repos, couché |
0.092 |
|
|
Serrurerie |
0.238 |
|
|
Ski, neige damée sur colline, vitesse maximale |
1.147 |
|
|
-sur le plat, marche |
0.599 |
|
|
-sur le plat à vitesse modérée |
0.498 |
|
|
Ski, neige poudreuse -loisir (F) |
0.465 |
|
|
-loisir (H) |
0.410 |
|
|
Squash |
0.887 |
|
|
Sylviculture, abattre des arbres |
0.552 |
|
|
-corder du bois |
0.356 |
|
|
-couper à la hache, lent |
0.356 |
|
|
-idem, rapide |
1.243 |
|
|
-désherber |
0.301 |
|
|
-écorcer des arbres |
0.515 |
|
|
-émonder des arbres |
0.540 |
|
|
-planter à la main |
0.456 |
|
|
-sarcler |
0.381 |
|
|
-scier machine |
0.314 |
|
|
-scier à la main |
0.511 |
|
|
-transporter billots |
0.778 |
|
|
Tennis |
0.456 |
|
|
Tennis de table |
0.285 |
|
|
Tir à l'arc |
0.272 |
|
|
Travail à la ferme conduire moissonne |
0.167 |
|
|
-conduire tracteur |
0.155 |
|
|
-enfourcher des balles de foin |
0.578 |
|
|
-nettoyer l'étable |
0.565 |
|
|
-nourrir le bétail |
0.356 |
|
|
-nourrir les animaux |
0.272 |
|
|
-pelleter du grain |
0.356 |
|
|
-traire à la machine |
0.096 |
|
|
-traire à la main |
0.226 |
|
|
Travail d'électricité |
0.243 |
|
|
Tricot, couture (H) |
0.096 |
|
|
Tricot, couture (F) |
0.092 |
|
|
Volley-ball |
0.209 |
|
|
OCCUPATION |
moyenne |
minimum |
maximum |
||
|
HOMMES (Kj) |
|
|
|
||
|
Retraités |
9 750 |
7 320 |
11 760 |
||
|
Cols blancs |
10 550 |
7 620 |
13 690 |
||
|
Commis de bureau |
11 720 |
9 750 |
13 770 |
||
|
Techniciens labo |
11 890 |
9 380 |
15 990 |
||
|
Travailleurs industriels âgés |
11 890 |
9 120 |
15 530 |
||
|
Etudiants université |
12 260 |
9 500 |
18 460 |
||
|
Travailleurs construction |
12 560 |
10 210 |
15 610 |
||
|
Travailleurs sidérurgie |
13 730 |
10 880 |
16 570 |
||
|
Cadets de l'armée |
14 610 |
12 510 |
17 160 |
||
|
Paysans Suisses âgés |
14 770 |
9 250 |
20 930 |
||
|
Fermiers |
14 860 |
10 250 |
19 550 |
||
|
Mineurs de charbon |
15 320 |
12 430 |
19 090 |
||
|
Travailleurs de la forêt |
15 360 |
11 970 |
19 250 |
||
|
FEMMES (Kj) |
|
||||
|
Ménagères âgées |
8 330 |
6 240 |
10 090 |
||
|
Ménagères âge moyen |
8 750 |
7 370 |
9 710 |
||
|
Assistantes labo |
8 910 |
5 610 |
10 630 |
||
|
Employées de magasin |
9 420 |
7 620 |
11 930 |
||
|
Etudiantes université |
9 580 |
8 750 |
10 460 |
||
|
Employées manufacture |
9 710 |
8 240 |
12 470 |
||
|
Employées pâtisserie |
10 510 |
8 290 |
14 190 |
||
|
Paysannes Suisses âgées |
12 100 |
9 210 |
16 160 |
||
La dépense énergétique est
obtenue en multipliant le nombre de minutes consacrées à l'activité par
l'équivalent énergétique de l'activité considérée.
Dépense
énergétique approximative des activités manuelles les plus courantes.
(D'après les données de E.W.
Bannister et S.R. Brown et P.-C. Goepfert et J.-C. Chignon)
Activités professionnelles et de loisirs METS KJ(min.kg)
TRES LEGERE , < 3
TRAVAIL ASSIS
Travail de bureau,
dactylographie, assemblage
0,113 à 0,130
et réparation électronique,
mécanique de précision.
0,105 à 0,150
TRAVAIL EN STATION DEBOUT
Vendeur, barman, portier.
0,14
CONDUITE
de camion, tracteur, machine
agricole, grue, engins divers.
0,16
TRAVAUX MENAGERS LEGERS
Couture, tricot, repassage
épluchage, laver la vaisselle,
0,092 à 0,188
mettre et débarrasser la
table, petit lavage de vêtement.
0,120 à 0,160
JARDINAGE LEGER
Taille de rosiers,
d'arbustes, tonte gazon sur tracteur, semer,
0,120 à 0,160
utiliser des outils à manche
court (plantoir, truelle).
0,16
LEGERE
3 à 7
POSTES DE L'INDUSTRIE
MECANIOUE
ET METALLURGlQUE
Ajustage, montage, travail
sur machine-outil.
0,15 à 0,243
Travail à la chaîne à
cadence et charges moyennes.
0,150 à 0,200
TRAVAIL DE GARAGE
Réparation auto, changement
de roue, lavage de voiture, camion.
0,200 à 0,260
CONDUITE
Camion remorque avec
manœuvres et sorties fréquentes
0,260 à 0,280
MAGASINAGE
Rangement et port d'objets
de masse <
sur des étagères à
mi-hauteur.
0,150 à 0,251
MENUISERIE LEGERE
0,22
INSTALLATION ELECTRIOUE
0,24
CONSTRUCTION D'UN MUR
0,30
Pose de pierres et briques,
mélange mécanique du ciment.
0,26
BRICOLAGE MODERE
Fendre du petit bois,
peinture intérieure, pose de papier,
0,142 à 0,201
Vissage (plus coûteux bras
en l'air).
0,112 à 0,180
JARDINAGE MODERE
Sarcler, biner, tailler
haies, arbustes, usage
0,200 à 0,350
tronçonneuse légère,
usage tondeuse tractée,
pousser une brouette <
TRAVAUX MENAGERS MODERES
Utilisation d'instruments à
manche long: aspirateur,
0,200 à 0,300
brosse à tapis
balai mécanique, passer la
serpillière,
0,26
laver les carreaux, murs,
placards
0,243 à 0,260
faire les lits, ranger une
pièce,
0,110 à 0,250
faire les courses <
MOYENNE
5 à 7
TRAVAUX DE BATIMENT
Menuiserie lourde,
charpente, réfection extérieure,
0,300 à 0,400
construction
maison travail de plâtrier,
peinture extérieure, brouettage >
Montage charpente
métallique, maniement du marteau
0,400 à 0,500
pneumatique, pelletage de
terre légère, grain, ordure,
0,347 à 0,360
lentement
PORT DE CHARGE
BRICOLAGE
Scier du bois tendre, fendre
à la hache
0,314 à 0,511
MONTER LES ESCALIERS
LENTEMENT
0,200 à 0,300
JARDINAGE
Bêchage terre légère,
tondeuse à traction manuelle en terrain
0,300 à 0,400
horizontal, ou petit
motoculteur.
0,300 à 0,400
Activités professionnelles et de loisirs METS KJ (mm.kg)
LOURDS 7
à 9
MANIEMENT D'OUTILS LOURDS
Piochage, creuser un trou et
terrassement divers, barre
0,393 à 0,452
masse, tronçonneuse lourde.
0,300 à 0,400
ENTRETIEN DE FOURS
INDUSTRIELS 0,53
PORTER
Charger un camion, fourcher
des bottes de foin,
0,400 à 0,578
déplacements de poids de
BRICOLAGE
Scier du bois dur, raboter
du bois tendre.
0,51
JARDINAGE
Bêchage lourd, arrachage
rapide des pommes de terre,
0,35
pelletage lourd (
TRES LOURD >
à 9
BUCHERON
0,500 à 1,243
MANŒUVRE LOURDE
Fourcher des balles de
paille
0,58
Raboter du bois dur
0,400 à 0,600
Fauchage rapide
0,50
Pelletées lourdes (
Porter des charges >
Porter des charges >
REMAROUES: 1 MET est la dépense
métabolique de base par kg et par minute d'un sujet éveillé, au repos et assis.
1 MET = 3,5 ml O2/kg/mn. 1 cal = 4,185 joules.
Tenir compte que le travail ménager
comporte des moments de travail physique intense.
Limites d'ordre
quantitatif pour le transport d'une charge.
Transport manuel pour un homme:
FxDxP<800 F <
D <
P < 10 T/j
F = Charge transportée ou manutentionnée en
kilos
P = Production journalière en Tonnes/Jour
D = Distance parcourue avec la charge
Manutention de charges au poste de
travail (déplacement <
Homme: P<14;
Femme: P<8;
(Valeurs tirées de la revue
"Travail et sécurité", 462, 1991, reprenant les travaux de l'INRS,
165, 1994 et du laboratoire de physiologie du travail).
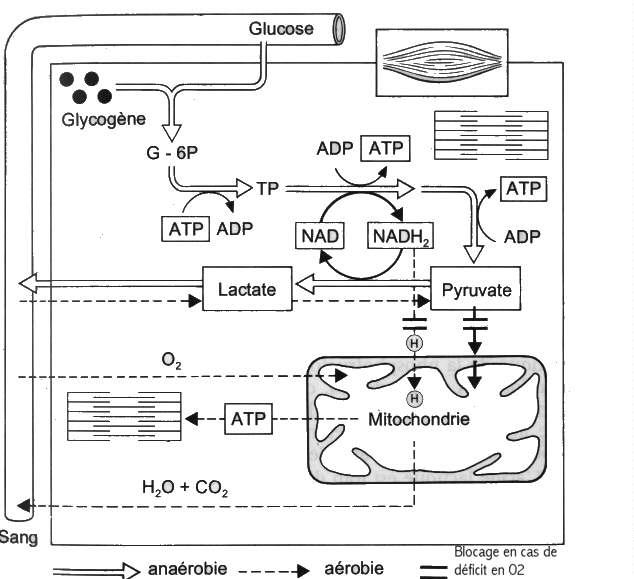
Pour assumer toutes ses fonctions, le corps
a besoin d'énergie. Cette énergie est constituée par la rupture des liaisons de
la molécule adénosine triphosphate (ATP). L'ATP est donc le carburant utilisé par
tous les processus du corps.
La production d'énergie se classe en deux
cycles principaux :
sans utilisation d'oxygène (anaérobie)
avec utilisation d'oxygène (aérobie)
Les processus anaérobies ont lieu dans le cytoplasme
de la cellule tandis que les processus aérobies ont lieu dans les
mitochondries.
Le réticulum endoplasmique ou sarcoplasmique
(cellule musculaire) constituent entre autre le site de la synthèse des
protéines.


1°) La glycolyse anaérobie
alactique: ATP et CP
 L'organisme trouve sa première source énergétique dans l'ATP présent
dans le muscle qui lui donne deux à trois secondes d'énergie. A aucun moment,
l’organisme n'emmagasine plus de
L'organisme trouve sa première source énergétique dans l'ATP présent
dans le muscle qui lui donne deux à trois secondes d'énergie. A aucun moment,
l’organisme n'emmagasine plus de
L'organisme puise ensuite dans la créatine
phosphate (CP) disponible dans les cellules musculaires pour obtenir une
énergie durable jusqu'à approximativement la dixième ou quinzième seconde (en
fonction notamment du degré d'entraînement du sujet). La concentration de C.P.
est environ de 3 à 5 fois celle de l’A.T.P. C’est pourquoi la CP est considérée
comme le réservoir de phosphates à haute énergie.
Il faut environ 29 Kj (7 Kcal) pour
synthétiser une mole d’A.T.P. L'efficacité relative du processus de transport
d’électrons (phosphorylation oxydative) pour retenir l’énergie chimique, est
approximativement de 40%. Le reste de l'énergie, 60%, est transformé en chaleur
dans l’organisme.
Chaque kilogramme de muscle contient environ
5 millimoles (mmol) d’A.T.P. et 15
mmol de C.P. La resynthèse des phosphagènes,
l'exercice fini, atteint 70% en 30 s (ATP +
CP). Elle est terminée dans un délai de 3 à 5 minutes.

2°) La glycolyse anaérobie lactique:
le glycogène et l'acide lactique

Les réserves de phosphate épuisées,
l'organisme se tourne vers les nutriments pour trouver dans leur dégradation
l'énergie nécessaire. Le premier appel est fait au glucose.
Les glucides sont les seuls aliments capable
de fournir l'énergie nécessaire à la formation de l'ATP en anaérobie. La
dégradation des glucides fournit en sus des éléments indispensables au
métabolisme des lipides et des protéines. La dégradation du glucose pour
fournir de l'ATP s'appelle
L'énergie délivrée de manière anaérobique
sera utilisée préférentiellement par les fibres rapides, l'énergie libérée de
manière aérobique sera utilisée préférentiellement par les fibres lentes
puisqu'elles possèdent de nombreuses mitochondries.
L’ensemble des réactions de glycolyse
anaérobie est pleinement efficace entre la 30ème et la 120ème
seconde, est optimal pendant 60 s pour disparaître vers la 180ème seconde.
Le mécanisme de la glycolyse n'est possible
qu'en présence du coenzyme NAD.



La dégradation du glycogène et du glucose
musculaire sans oxygène prend donc ensuite le relais de ATP+CP. Cette énergie
est productrice d'acide lactique et donc d'ions H+. De ce fait, les cellules
musculaires cesseront de fonctionner quand l'accumulation d'H+ deviendra trop
importante. Les cellules musculaires peuvent aussi cesser de fonctionner quand
le taux d'usure du glycogène devient trop important. En effet l'évolution
naturelle a doté l'organisme d'un contrôle du glycogène afin de garder toujours
une réserve indispensable pour faire face à une situation vitale et donc à
l'éternel choix de fuir ou combattre; ces actions vitales nécessitent une
source d'énergie immédiatement disponible, qui sont ATP + CP et le glycogène
assurant la synthèse rapide de l'ATP. Le dopage ou l'extrême motivation
permettent de puiser dans cette réserve vitale. D'autre part le catabolisme du
glycogène fournit des substrats indispensables à l'utilisation aérobie du glucose
et des lipides. Ce fait est à l'origine de la formule "les lipides brûlent
au feu des glucides", qui valide la proposition suivant laquelle le
glycogène doit être épargné pour permettre un effort de longue durée. Une
intensité de départ trop importante d'un exercice, nuit donc à sa longévité.




Le glycogène est le polysaccharide
synthétisé à partir du glucose lors de la glycogenèse et emmagasiné dans les
tissus animaux. Les molécules de glycogène sont habituellement volumineuses.
Leur taille varie de quelques centaines à quelques milliers de molécules de
glucose attachées entre elles. L’organisme d’êtres humains convenablement
nourris, contient environ 375 à
C'est la quantité d'oxygène disponible dans
la cellule qui détermine le type de processus utilisé. En début d'exercice,
l'insuffisance d'O2 dans la cellule impose la glycogénolyse comme
processus, le temps que la cellule fasse le plein d'O2.
La filière du glycogène est disponible après
quelques secondes (5 s), atteint son maximum de puissance entre la première et
la deuxième minute, le plus souvent vers la 20ème seconde, avant que
le système d'économie du glycogène n'intervienne pour progressivement
l'épargner au profit des acides gras libres.
Au delà d'un effort de 50% de VO2
Max pour un sujet non entraîné et 60 à 70% pour un sujet entraîné la
participation de la filière anaérobique lactique devient plus importante et
croît avec l'intensité de l'effort au delà de V02 Max. Les fibres
rapides consommatrices de glycogène sont progressivement recrutées. Le seuil
anaérobique déterminera le moment ou l'organisme produit plus d'acide lactique
que ne peut en tamponner l'oxygène acquis (seuil d'accumulation sanguine
d'acide lactique). Ce seuil passé, le sujet va s'épuiser et cesser l'effort
entre 3 et 15 minutes. Le seuil anaérobique peut être au delà de 90% de VO2
Max pour de très grands sportifs en endurance.

£ Pendant l’exercice physique, les hydrates de carbone
mis en réserve sous forme de glycogène musculaire servent de source d’énergie
au muscle même qui les renferme. Le glycogène stocké dans un muscle n'est
utilisable que par ce muscle, il ne peut-être transféré d'un muscle à l'autre.
£ Dans le foie, le glycogène est reconverti en glucose,
puis transporté dans le sang pour satisfaire les éventuels besoins des muscles
sollicités. Le mot glycogénolyse réfère à ce processus de conversion,
méthode de fourniture rapide de glucose aux muscles impliqués dans toute forme
de travail. C'est l'augmentation du calcium cytoplasmique pendant la
contraction musculaire qui est à l'origine de la glycogénolyse.
£ Les réserves de glucose sont de
£ Lorsque les réserves de glycogène sont épuisées à la
suite d’une restriction alimentaire ou d’un exercice physique, la synthèse du
glucose à partir des composantes structurales des autres nutriments, et
particulièrement les protéines, tend à augmenter. Ce processus constitue la néoglucogenèse.
Le glycogène augmente la teneur en potassium
et donc la conduction nerveuse et la contractilité musculaire.
Une concentration normale de glycogène
atteint 15 g/kg de muscle, une surcompensation permet d'atteindre
L'insuffisance musculaire en glycogène n'a
pas d'influence sur l'intensité de l'exercice mais sur
Le glycogène est le seul nutriment qui peut
être catabolisé en anaérobiose pour la resynthèse d’A.T.P.; ce substrat est de
toute évidence hautement prioritaire dans le "moulin métabolique"; au
cours d’un exercice vigoureux, il semble que ce soit l’intensité de l’effort
qui détermine l’épuisement spécifique des fibres rapides ou celui des fibres
lentes.
Les hormones, spécialement l’insuline,
jouent un rôle important dans la régulation des réserves hépatiques et
musculaires de glycogène en contrôlant le niveau de sucre sanguin en circulation.
L'insuffisance de glycogène hépatique peut entraîner des signes d'hypoglycémie
alors même que le stock de glycogène musculaire n'est pas épuisé (en général
dans des exercices de longue durée d'intensité modérée).
60% du glucose hépatique est destiné aux
besoins du SNC dont le métabolisme ne consomme que du glucose pur. Ainsi, une
personne faisant une grève de la faim, doit ingérer un minimum de glucose
journalier sous peine de sombrer rapidement dans le coma. La participation du
glycogène musculaire au maintien de la glycémie se fait d’une manière indirecte
par le cycle de Cori: glycogène musculaire à acide lactique qui passe du
muscle dans le sang et parvient au foie où il peut donner du glucose.
 Le processus le plus efficace pour augmenter les réserves musculaires en
glycogène comprend des exercices continus intenses destinés à vider les muscles
de leur glycogène, associés pendant plusieurs jours à un régime hypoglucidique,
suivi de repos associé à un régime hyperglucidique. Les réserves de glycogène
musculaire peuvent ainsi dépasser
Le processus le plus efficace pour augmenter les réserves musculaires en
glycogène comprend des exercices continus intenses destinés à vider les muscles
de leur glycogène, associés pendant plusieurs jours à un régime hypoglucidique,
suivi de repos associé à un régime hyperglucidique. Les réserves de glycogène
musculaire peuvent ainsi dépasser




La surcompensation des réserves de glycogène
n’est pas conseillée dans les sports comme le sprint, car l’eau retenue par le
glycogène alourdit le sujet.

 Au repos la concentration plasmatique d’acide lactique = 10 mg/100 ml de
sang
Au repos la concentration plasmatique d’acide lactique = 10 mg/100 ml de
sang
La plus forte et la plus rapide concentration
d’acide lactique est enregistrée après un exercice soutenu de 60 à 180
secondes. A environ 50% de la capacité maximale du métabolisme aérobie d’un
sujet en santé non entraîné, l’acide lactique commence à s’accumuler. En
moyenne, chez l’adulte sédentaire, la lactatémie sanguine n’augmente pas pour
des efforts d’intensité faible, correspondant à moins de 50% du VO2
Max. Les lactates apparus en début d’exercice disparaissent progressivement. La
courbe présente une partie horizontale. Pour des intensités plus fortes (50 à
70% du VO2 Max) le taux de lactates sanguins augmente et se
stabilise en quelques minutes, modifiant ainsi la pente de

Le lactate produit par le muscle n'est que
partiellement libéré dans le sang, ce qui réduit l'acidose. Les réactions
aérobies fournissent l’énergie nécessaire à l’effort et l’acide lactique
produit est soit oxydé. soit reconverti en glucose, probablement au niveau du
foie et peut-être des reins. En régime stable, l’accumulation d’acide lactique
est minimale.
En apparence, la majeure partie de l’acide
lactique est oxydée pour en libérer l'énergie. En fait, on sait très bien
maintenant que le cœur, le foie, les reins et les muscles squelettiques
utilisent l’acide lactique plasmatique comme substrat énergétique aussi bien
pendant l’exercice que pendant la récupération.
L’acide lactique se dégrade en acide
pyruvique, produit final de la glycolyse qui entre ensuite dans le cycle de
Krebs, en présence d’oxygène (glycolyse aérobie). Sans oxygène (glycolyse
anaérobie), l'acide pyruvique est reconverti en acide lactique sans autre
resynthèse d'ATP (néoglucogenèse).
Le cycle de Cori (l'acide lactique redevient
acide pyruvique) n’est pas seulement un refuge pour l’acide lactique mais aussi
un moyen d'augmenter le glucose sanguin et le glycogène musculaire.
 Hypothèses de la diminution d’acide lactique dans un
exercice sous-maximal
Hypothèses de la diminution d’acide lactique dans un
exercice sous-maximal
a) Une plus
grande utilisation des acides gras entraîne une moins grande utilisation de
glycogène et, par conséquent, une moins grande production d’acide lactique.
b) La dette
d’oxygène contractée au début de l’exercice pourrait être plus petite à cause
d’une augmentation rapide de la consommation d’oxygène. Ceci pourrait aussi
provoquer une moins grande accumulation d’acide lactique; d'où la nécessité de
s'échauffer et de laisser le temps à la consommation d'O2 de se
stabiliser pour pouvoir durer et bénéficier d'un exercice plus facile à
supporter. Plus la dette d'O2 de départ est grande plus l'exercice
est ressenti comme difficile.

3°) La glycolyse aérobie: les acides
gras libres
 La glycolyse anaérobie a transformé le glucose en acide pyruvique, et
l'organisme va tirer de cet acide pyruvique les 95% d'énergie restant
disponible par un mécanisme très important appelé le cycle de Krebs. Le
cycle de Krebs assure la transformation aérobique de l'acide pyruvique, des
lipides et des acides aminés en ATP. Cette transformation est assurée à
l'intérieur des mitochondries qui tiennent le rôle de centrale énergétique.
La glycolyse anaérobie a transformé le glucose en acide pyruvique, et
l'organisme va tirer de cet acide pyruvique les 95% d'énergie restant
disponible par un mécanisme très important appelé le cycle de Krebs. Le
cycle de Krebs assure la transformation aérobique de l'acide pyruvique, des
lipides et des acides aminés en ATP. Cette transformation est assurée à
l'intérieur des mitochondries qui tiennent le rôle de centrale énergétique.
 La troisième filière fait appel à la dégradation du glucose, de l'acide
lactique, de certains acides aminés (alanine), et surtout des acides gras
libres en présence d'oxygène. Il y a glycolyse aérobie du glucose (glycogène)
et glycolyse aérobie des lipides.
La troisième filière fait appel à la dégradation du glucose, de l'acide
lactique, de certains acides aminés (alanine), et surtout des acides gras
libres en présence d'oxygène. Il y a glycolyse aérobie du glucose (glycogène)
et glycolyse aérobie des lipides.
La glycolyse aérobie du glucose produit de
l’énergie et de l’acide pyruvique qui est ensuite catabolisé dans le cycle de
Krebs. L'utilisation aérobie du glycogène produit 13 fois plus d'énergie que
son utilisation anaérobie.
 La disponibilité sans limite des acides gras libres dans les réserves
adipeuses, fait que la limitation de cette filière sera la consommation
maximale de l'oxygène (VO2 Max). La bonne oxygénation du muscle est
d'autant plus indispensable pour pouvoir utiliser les substrats lipidiques. Les
muscles vont utiliser en présence d'oxygène les cellules lipidiques qu'ils
contiennent et les acides gras présents dans le sang. Ces réserves sont rapidement
épuisées et le muscle va faire appel aux réserves lipidiques du tissu adipeux.
Les adipocytes vont donc libérer les acides gras et le glycérol par la lipolyse
et le plasma va véhiculer les AGNE (acides gras non esthérifiés) au muscle. Les
AGNE pénètrent ensuite dans le cytoplasme de la cellule, et enfin dans la
mitochondrie sous l'action de la carnitine. les AGNE seront ensuite dégradés en
acétyl-coenzyme A et pris en charge par le cycle de Krebs pour produire de
l'ATP.
La disponibilité sans limite des acides gras libres dans les réserves
adipeuses, fait que la limitation de cette filière sera la consommation
maximale de l'oxygène (VO2 Max). La bonne oxygénation du muscle est
d'autant plus indispensable pour pouvoir utiliser les substrats lipidiques. Les
muscles vont utiliser en présence d'oxygène les cellules lipidiques qu'ils
contiennent et les acides gras présents dans le sang. Ces réserves sont rapidement
épuisées et le muscle va faire appel aux réserves lipidiques du tissu adipeux.
Les adipocytes vont donc libérer les acides gras et le glycérol par la lipolyse
et le plasma va véhiculer les AGNE (acides gras non esthérifiés) au muscle. Les
AGNE pénètrent ensuite dans le cytoplasme de la cellule, et enfin dans la
mitochondrie sous l'action de la carnitine. les AGNE seront ensuite dégradés en
acétyl-coenzyme A et pris en charge par le cycle de Krebs pour produire de
l'ATP.
Ces substrats ne produisent pas d'énergie
mais nécessitent la présence de glycogène pour être utilisés. En effet, la
dégradation des acides gras semble dépendre du fonctionnement continuel du
catabolisme du glucose ("les lipides brûlent au feu des glucides").
Il faut se rappeler que l’acétyl-CoA, entre dans le cycle de Krebs en se
combinant à l'acide oxalo-acétique (obtenu surtout par le catabolisme des
sucres) pour former l’acide citrique. La dégradation des acides gras dans la
mitochondrie, au cours du cycle de Krebs, se poursuit tant qu’une quantité
suffisante d’acide oxalo-acétique peut se combiner à l’acétyl-CoA, sous-produit
de
 L‘acide pyruvique formé au cours du métabolisme du glucose pourrait être
une source importante d’acide oxalo-acétique. Comme la conversion de l’acide
pyruvique en acétyl-CoA est irréversible, les acides gras ne peuvent pas être
utilisés pour refaire le glucose. L'insuffisance de glucose dans des situations
d'endurance extrêmes produit une accumulation de fragments d'acétate, formés
durant la bêta-oxydation des acides gras. Si ces fragments d'acétate ne peuvent
rentrer dans le cycle de Krebs, ils sont transformés en corps cétoniques qui se
retrouvent dans l'urée et dans le sang.
L‘acide pyruvique formé au cours du métabolisme du glucose pourrait être
une source importante d’acide oxalo-acétique. Comme la conversion de l’acide
pyruvique en acétyl-CoA est irréversible, les acides gras ne peuvent pas être
utilisés pour refaire le glucose. L'insuffisance de glucose dans des situations
d'endurance extrêmes produit une accumulation de fragments d'acétate, formés
durant la bêta-oxydation des acides gras. Si ces fragments d'acétate ne peuvent
rentrer dans le cycle de Krebs, ils sont transformés en corps cétoniques qui se
retrouvent dans l'urée et dans le sang.
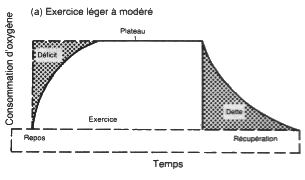
 Le temps nécessaire pour atteindre le maximum des réactions aérobies
varie de 1 minute chez le sportif entraîné à 4 minutes chez le sédentaire. Le
substrat lipidique est sollicité progressivement dès la 10ème
seconde d'effort, la puissance maximale de la glycolyse aérobie est atteinte
vers la 45ème seconde, mais son utilisation ne devient intéressante
qu'entre la 5ème et 10ème minute d'effort. acétyl-CoA
provenant des AGL entre en compétition avec celui provenant du pyruvate et
réduit ainsi la consommation de glycogène. Plus l'effort va se prolonger, plus
le substrat lipidique va être sollicité pour économiser le glycogène, notamment
à l'épuisement du glycogène hépatique, sollicité depuis la 10ème ou
15ème minute; au bout de 2 h 30 à 3 heures d'exercice à 70% de VO2
Max, les voies lipolytiques sont utilisées à leur maximum. Arrivé à un certain
stade de puissance (50% VO2 Max pour un sujet non entraîné et 60 à
70% pour un sujet entraîné) la puissance supplémentaire sollicitée par
l'exercice sera assurée par la filière anaérobique lactique et les fibres
rapides.
Le temps nécessaire pour atteindre le maximum des réactions aérobies
varie de 1 minute chez le sportif entraîné à 4 minutes chez le sédentaire. Le
substrat lipidique est sollicité progressivement dès la 10ème
seconde d'effort, la puissance maximale de la glycolyse aérobie est atteinte
vers la 45ème seconde, mais son utilisation ne devient intéressante
qu'entre la 5ème et 10ème minute d'effort. acétyl-CoA
provenant des AGL entre en compétition avec celui provenant du pyruvate et
réduit ainsi la consommation de glycogène. Plus l'effort va se prolonger, plus
le substrat lipidique va être sollicité pour économiser le glycogène, notamment
à l'épuisement du glycogène hépatique, sollicité depuis la 10ème ou
15ème minute; au bout de 2 h 30 à 3 heures d'exercice à 70% de VO2
Max, les voies lipolytiques sont utilisées à leur maximum. Arrivé à un certain
stade de puissance (50% VO2 Max pour un sujet non entraîné et 60 à
70% pour un sujet entraîné) la puissance supplémentaire sollicitée par
l'exercice sera assurée par la filière anaérobique lactique et les fibres
rapides.
L'entraînement permet d'améliorer le temps
de sollicitation des lipides et leur part énergétique de l'effort. L'organisme
apprend à économiser plus rapidement le précieux glycogène en transférant la demande
énergétique vers les acides gras.
La vitesse de renouvellement des AGL va de 3
minutes au repos à 20 secondes à l'exercice. Le métabolisme des graisses est
plus important dans un muscle entraîné.
La consommation d'oxygène nécessaire à la
production aérobique de l'énergie va augmenter proportionnellement à l'effort
et à la production d'énergie jusqu'au stade ou l'organisme ne peut plus
apporter suffisamment d'oxygène et va devoir faire appel au mécanisme
anaérobique de production d'énergie, ce qui assurera près de 60 secondes
d'effort maximal disponible. En fait lorsqu'on atteint 50 à 55% de la
consommation maximale d'oxygène, les mécanismes anaérobiques commencent à
dépasser les mécanismes aérobiques et à stocker de l'acide lactique, puisqu'il
n'y a plus assez d'oxygène pour l'utiliser. L'acide lactique va être apporté
aux muscles par la voie circulatoire et va installer la sensation de fatigue
obligeant l'organisme à arrêter l'exercice. En fait les ions H+ hydrogène de
l’acide lactique ne trouvent plus assez d’ions O oxygène pour se combiner en
eau H2O. Les cellules vont donc arrêter de fonctionner pour stopper
le mécanisme d’accumulation des ions H+ (acidose), mortel à terme.
La perte des minéraux et de l'eau par la
sueur est également une cause d'arrêt de l'exercice par fatigue ou crampes
musculaires. L'acide lactique est ensuite excrété ou resynthétisé en acide
pyruvique et en énergie disponible si il y a suffisamment d'oxygène pendant la
phase de récupération.
Le débit cardiaque est le principal indicateur
des possibilités aérobiques du sujet. Le débit cardiaque est le produit de la
fréquence cardiaque par le volume systolique exprimé en litres par minute. Plus
le débit sanguin augmente, plus la concentration d'AGL disponible et son
utilisation augmente. Les fibres musculaires à secousses lentes, au débit
sanguin volumineux, aux nombreuses mitochondries, constituent le site idéal de
dégradation des graisses. Le métabolisme lipidique est freiné par l'acide
lactique; cependant dès que le taux de lactates a diminué la mobilisation des
AGL peut intervenir.
D'après ASTRAND, les lipides participeraient
plus à la fourniture d'énergie dans un exercice intermittent que dans un
exercice continu, mais cette donnée n'est pas retrouvée chez les autres
auteurs.
4°) la filière protéique: la
néoglucogenèse
 Il n'existe pas de réserves énergétiques de protéines. Cependant il
existe un mécanisme d'utilisation des protéines qu'on retrouve essentiellement
à la fin des efforts aérobiques de très longue durée et qui consiste à détruire
l'alanine musculaire pour la transformer en glucose, toujours dans le but
d'économiser le précieux glycogène musculaire. Ce mécanisme qui consiste à
détruire les muscles pour alimenter le SNC en glucose se retrouve dans les
grèves de la faim, ou dans les régimes hypoglucidiques aberrants.
Il n'existe pas de réserves énergétiques de protéines. Cependant il
existe un mécanisme d'utilisation des protéines qu'on retrouve essentiellement
à la fin des efforts aérobiques de très longue durée et qui consiste à détruire
l'alanine musculaire pour la transformer en glucose, toujours dans le but
d'économiser le précieux glycogène musculaire. Ce mécanisme qui consiste à
détruire les muscles pour alimenter le SNC en glucose se retrouve dans les
grèves de la faim, ou dans les régimes hypoglucidiques aberrants.
La néoglucogenèse recycle les substrats
provenant de la lipolyse (glycérol) de la protéolyse (alanine) et de la
glycolyse (lactate). La dégradation des protéines en glucose pendant l'exercice
entraîne une augmentation de l'urée sanguine (acide urique).
Bien que la principale région de
désamination (extraction de l’azote) soit le foie, les muscles squelettiques
contiennent aussi des enzymes qui retirent l’azote des acides aminés pour le
transférer à d’autres composés par un processus de transamination. L’alanine,
par exemple, perd son amine mais gagne un oxygène avec une double liaison,
formant ainsi l’acide pyruvique. L‘acide glutamique se transforme en acide
alpha-cétoglutamique et l’acide aspartique en acide oxalo-acétique; tous ces
produits appartiennent au cycle de Krebs. Le foie utilise le lactate et le
pyruvate pour produire du glucose par l'intermédiaire du cycle de Cori.
 Il y a utilisation des protéines dès le début de l'exercice (15 à 20 s),
ce qu'indique l'augmentation de la concentration en azote intramusculaire. Une
partie de l'azote se combine avec le pyruvate qui n'a pas intégré le cycle de
Krebs pour produire de l'alanine. Ceci explique l'augmentation des synthèses
protéiques observées dès la fin de l'exercice. Le rôle des protéines dans
l'approvisionnement en glucose est important pour les exercices prolongés
supérieurs à 3 h, car son action augmente avec la durée de l'effort au-delà de
2 h. Le rôle néoglucogénique du glycérol est particulièrement important lorsque
l’apport alimentaire de glucides est limité au cours d’un exercice prolongé qui
vide substantiellement les réserves de glycogène.
Il y a utilisation des protéines dès le début de l'exercice (15 à 20 s),
ce qu'indique l'augmentation de la concentration en azote intramusculaire. Une
partie de l'azote se combine avec le pyruvate qui n'a pas intégré le cycle de
Krebs pour produire de l'alanine. Ceci explique l'augmentation des synthèses
protéiques observées dès la fin de l'exercice. Le rôle des protéines dans
l'approvisionnement en glucose est important pour les exercices prolongés
supérieurs à 3 h, car son action augmente avec la durée de l'effort au-delà de
2 h. Le rôle néoglucogénique du glycérol est particulièrement important lorsque
l’apport alimentaire de glucides est limité au cours d’un exercice prolongé qui
vide substantiellement les réserves de glycogène.
Le cycle de Cori est un moteur important de
la fourniture d'énergie quand il se passe plus de 5 heures entre les repas,
d'où la nécessité de repas régulièrement espacés pour que le maintien de la
glycémie ne soit pas assuré par la destruction des protéines.
Les quatre filières énergétiques
fonctionnent ensemble avec des temporisations propres à des degrés de participation
différents.
Le glycérol provient du glucose ou du
pyruvate en l'absence de glucose ou des pentoses.
Le catabolisme des glucides revêt quatre
possibilités:
ê
voie d'Embden-Meyerhoff ou voie énergétique de l'activité: le glucose
est conduit jusqu'au cycle de Krebs;
Þ
voie de la glycogénogenèse: une partie peu importante de glucose est
stockée en glycogène dans le foie (75 à
ò
voie de la lipogenèse physiologique: fourniture du glycérol activé
utilisé par les adipocytes pour stocker les triglycérides (utilisation des
trioses-phosphates par l'activité catalytique de NAD et NADH);
¯ voie
des pentose-phosphates ou voie de Dickens-Horecker: la dégradation du
glucose6-phosphate aboutit à la formation de divers pentoses-phosphates et de
NADP qui intervient dans la biosynthèse des acides gras et des stéroïdes. La
voie des pentoses est favorisée par une alimentation riche en glucose, une
prise alimentaire journalière non fractionnée (1 repas). La quantité de glucose
prenant la voie des pentoses au lieu de la voie de Embden-Meyerhoff peut-être
décuplée par une seule prise alimentaire quotidienne. Cette voie provoque une
hypersécrétion d'insuline. Cette voie est la voie principale du métabolisme du
glucose chez l'obèse.
Fraction moyenne d'utilisation des
différents substrats pendant l'exercice
|
Processus |
Exercice maximal, durée du travail |
|||||||
|
|
10 s |
1 mn |
2 mn |
4 mn |
10 mn |
30 mn |
60 mn |
120 mn |
|
Anaérobie |
|
|||||||
|
Kcal |
25 |
40 |
45 |
45 |
35 |
30 |
20 |
15 |
|
% |
85 |
65-70 |
50 |
30 |
10-15 |
5 |
2 |
1 |
|
|
||||||||
|
Aérobie |
|
|||||||
|
Kcal |
5 |
20 |
45 |
100 |
250 |
700 |
1300 |
2400 |
|
% |
15 |
30-35 |
50 |
70 |
85-90 |
95 |
98 |
99 |
|
|
||||||||
|
Total |
|
|||||||
|
Kcal |
30 |
60 |
90 |
145 |
285 |
730 |
1320 |
2415 |
Substrats et rendements énergétiques maximum des
différentes sources énergétiques dans la cellule musculaire
 (R.GYM l'endurance de B. Villiger & autres aux éditions Masson)
(R.GYM l'endurance de B. Villiger & autres aux éditions Masson)
|
Source d'énergie |
Substrat |
Quantité de phosphates par Kg de muscle |
Durée de sollicitation maximum |
Rendement énergétique max µmol/g x s. |
|
Anaérobie alactique |
ATP |
env 6 mmol |
2 - 3 s |
|
|
|
CP |
env 20-25 mmol |
5 - 7 s |
|
|
|
Phosphate total |
env 30 mmol |
7 - 10 s |
1.6 - 3.0 |
|
Anaérobie lactique |
Glycogène |
env 270 mmol |
45 - 90 s |
1.0 |
|
Aérobie |
Glycogène |
env 3000 mmol |
45 - 90 mn |
0.50 |
|
|
Triglycérides |
env 50000 mmol |
plusieurs heures |
0.24 |
 Dans le cas d'un exercice bref et intense ne dépassant pas 30 minutes,
il y a :
Dans le cas d'un exercice bref et intense ne dépassant pas 30 minutes,
il y a :
¨ augmentation des catécholamines stimulant la
glycogénolyse hépatique
¨ augmentation du glucagon renforçant l'action des catécholamines
et entraînant une augmentation et donc une utilisation accrue des AGL
plasmatiques, même après l'exercice
¨ baisse de l'insuline stimulant la glycogenèse.
L‘égalité des contributions des processus
aérobies et anaérobies à la dépense totale d’énergie, est réalisée entre 2 et 4
minutes.
Au cours d’une course de
La filière aérobique intervient massivement
par rapport aux autres filières dans l’exercice musculaire durant 3 à 9
minutes; elle intervient presque exclusivement (90%) dans les exercices
musculaires durant plus de 10 minutes.
Durant un exercice sous-maximal:
¨ Le déficit d'oxygène se constitue pendant les trois
premières minutes; il est la conséquence de la régulation métabolique au niveau
musculaire.
¨ Lorsque l'exercice se prolonge l'utilisation de
glycogène musculaire chute au profit de l'utilisation de glucose sanguin.
¨ On note deux paliers importants dans la relation
glycogène/ AGL lors de l'exercice physique à 10 mn et 30 mn.
¨ Après 40 mn d'exercice, 75 à 90% du métabolisme
oxydatif est assuré par l'apport sanguin.
¨ Au bout
de 50 minutes, le cortisol augmente stimulant la néoglucogenèse hépatique.
¨ Le glycogène permet de tenir 80 minutes, le
glycogène hépatique donne 20 mn de plus, les acides gras donneront 100 minutes
supplémentaires et 1600 mn grâce aux acides aminés non structurels.
¨ A partir de 2 h 30 - 3 h, la testostérone baisse,
permettant la mobilisation des dernières réserves en glycogène, et augmentant
la mobilisation des réserves protéiques afin d'alimenter la néoglucogenèse.
 ¨ Pour
¨ Pour 

Rapport durée
de performance /voies énergétiques prédominantes
McArdle Wd, Katch Fl & Vl, exercice physiology, Lea & Febiger
1981)
 un effort modéré soutenu pendant 3 à 6 heures, le facteur limitant peut
être soit l’importance du stock de glycogène musculaire, soit l’abaissement de
Ia glycémie agissant surtout sur le système nerveux. On a démontré qu'un chien
sous perfusion glucosée aux carotides, augmentait son endurance, mettant ainsi
en évidence, la limitation due à la fatigue du SNC.
un effort modéré soutenu pendant 3 à 6 heures, le facteur limitant peut
être soit l’importance du stock de glycogène musculaire, soit l’abaissement de
Ia glycémie agissant surtout sur le système nerveux. On a démontré qu'un chien
sous perfusion glucosée aux carotides, augmentait son endurance, mettant ainsi
en évidence, la limitation due à la fatigue du SNC.

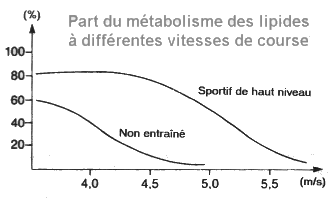

Le rapport entre l'utilisation d'hydrates de
carbone et de graisse est déterminé par le quotient respiratoire (QR = VO2
/ VCO2). La production de

Les mécanismes d'adaptation à
l'effort
Le volume d'éjection systolique
Il augmente linéairement jusqu'à la moitié
de l'intensité de l'effort (50% VO2 Max). Au delà, il se stabilise,
voir diminue, gêné par l'accélération du rythme cardiaque; la diminution du VES
quand le patient atteint presque VO2 Max (90%) signe la
prépondérance du travail anaérobique. On note des valeurs de 50 cm3
au repos et 120 cm3 à l'effort en moyenne. Ces valeurs peuvent être
augmentées de 50% par l'entraînement. Le volume systolique maximum est atteint
à 40-50% de VO2 Max. Si l'augmentation de VES par un sportif à
l'effort debout peut-être le double de la valeur de repos, à l'effort couché
elle ne sera que de 10 à 16%.
L'augmentation du VES dépend:
¢ de l'augmentation du volume télédiastolique du ventricule,
donc du volume sanguin total,
¢ de la compliance péricardique et myocardique,
¢ de la relaxation diastolique,
¢ du retour veineux,
¢ de la contractilité myocardique (sous contrôle
nerveux et hormonal), diminuant le volume télésystolique,
¢ des résistances périphériques (prépondérance de la
vasoconstriction a -adrénergique orthosympathique, facilitant le retour
veineux).
Le débit cardiaque est le produit de la
fréquence cardiaque par le volume d'éjection systolique. DC = Fc x VES.
Il augmente linéairement avec l'effort,
grâce à l'augmentation de VES et Fc jusqu'à 50% des capacités physiques. Au
delà, seule l'augmentation de Fc assure l'accroissement du débit cardiaque. Le
débit cardiaque de 4 à
 Chez le sujet sain, le rythme de repos du nœud sinusal est entre 90 et
110 bats/min, mais est freiné par le tonus parasympathique (tonus vagal). De ce
fait, chez le sujet moyen, le Fc de repos est de 70 bats/min et peut s'élever
jusqu'à 220 - âge ± 10.
Chez le sujet sain, le rythme de repos du nœud sinusal est entre 90 et
110 bats/min, mais est freiné par le tonus parasympathique (tonus vagal). De ce
fait, chez le sujet moyen, le Fc de repos est de 70 bats/min et peut s'élever
jusqu'à 220 - âge ± 10.
Les nerfs qui innervent le cœur et des substances chimiques en
circulation peuvent faire varier rapidement la fréquence cardiaque. 50% de
l'augmentation de Fc est due principalement à l'activité orthosympathique
notamment bêta, avec un relais important du ganglion stellaire droit, et à la
libération des neurotransmetteurs adrénaline et noradrénaline. Ces contrôles
extrinsèques peuvent accélérer le cœur par anticipation, avant même le début de
l’exercice. Dans une large mesure, ce contrôle extrinsèque peut abaisser la
fréquence cardiaque à 30 battements par minute chez des athlètes en endurance
bien entraînés, et l’élever jusqu’à 220 battements par minute au cours d’un
exercice maximal.

Ces valeurs sont grandement modifiables en
fonction de facteurs individuels (repos), externes (température) et surtout de l'indice
de sédentarité qui augmente Fc de repos et diminue Fc Max et de l'indice
d'activité qui exerce la modération inverse.
D’autres facteurs influencent la fréquence
cardiaque: émotion, apport alimentaire, posture, groupes musculaires
sollicités, modalité de travail (continu ou par intervalles) et type de
contraction musculaire (isométrique ou anisométrique). Par exemple, au cours
d’un travail exécuté par les membres supérieurs ou lors de contractions
musculaires isométriques (statiques), la fréquence cardiaque est beaucoup plus
élevée que si le travail sous-maximal était accompli par les membres
inférieurs. Par conséquent, si on utilisait la relation entre la fréquence
cardiaque et la consommation d’oxygène, établie au cours d’un exercice à
bicyclette ou à la course, pour estimer la consommation d’oxygène à partir
d’une fréquence cardiaque relevée au cours d’un exercice sollicitant le tronc
ou comportant des contractions isométriques, on obtiendrait une valeur
surestimée de la consommation d’oxygène.
Physiologie
de l'adaptation de Fc à l'effort
F Fc s'accélère avant même de débuter l'effort (commande sympathique).
F Fc augmente en 0.5 s, avec une évolution en dent de scie représentant la recherche du système nerveux autonome d'un nouvel équilibre.
F Après un premier seuil d'adaptation , Fc augmente linéairement à l'intensité de l'effort jusqu'au seuil anaérobique où elle augmente moins vite que l'intensité de l'effort.
F Par rapport à courbe de consommation maximale d'oxygène (VO2 Max), elle se ralentit à partir de 90% de VO2 Max.
n
Pour une même VO2, Fc est plus importante si la masse musculaire
engagée est plus petite.
n
Pour un même effort, plus VES augmente, moins Fc augmente.
n
A un même palier la Fc se stabilise progressivement en 2 à 3 minutes (steady
state). Cet équilibre est d'autant plus long à obtenir que les paliers sont
élevés.
n
La variation quotidienne de la fréquence cardiaque sous-maximale à la même
intensité de travail est d’environ 5 battements par minute.
 n Des fréquences cardiaques
anormalement basses peuvent indiquer un fonctionnement anormal du nœud
sino-auriculaire. L’impossibilité, pour la fréquence cardiaque, d‘augmenter à
l’effort, particulièrement lorsqu’elle s’accompagne de fatigue extrême, peut
aussi être un indice de stress cardiaque et de cardiopathie. L'incompétence
chronotropique est l'impossibilité de monter la Fc jusqu'à 95% de la Fc normale
à l'âge du sujet.
n Des fréquences cardiaques
anormalement basses peuvent indiquer un fonctionnement anormal du nœud
sino-auriculaire. L’impossibilité, pour la fréquence cardiaque, d‘augmenter à
l’effort, particulièrement lorsqu’elle s’accompagne de fatigue extrême, peut
aussi être un indice de stress cardiaque et de cardiopathie. L'incompétence
chronotropique est l'impossibilité de monter la Fc jusqu'à 95% de la Fc normale
à l'âge du sujet.
Le débit sanguin à l'effort du myocarde
le myocarde extrait en moyenne 60% à 70% de
l'oxygène (O2) du sang artériel coronarien (pour 10 à 20% au niveau
des autres organes). Cette capacité permet au myocarde de consommer 10% des
besoins en O2 de l'organisme alors que son débit sanguin à l'effort
n'est multiplié que par 3 à 5. Le débit sanguin du muscle à l'effort peut être
parallèlement multiplié par 10. La mise en jeu de masses musculaires
importantes est nécessaire pour entraîner la circulation centrale.
Le travail externe d'éjection systolique du
myocarde dépend du remplissage diastolique lié à l'importance du retour veineux
(pré-charge), et des résistances périphériques (post-charge) qui s'opposent à
l'écoulement du sang. Ce travail est caractérisé par la puissance de la
contraction.
II/ Adaptation de la distribution
sanguine régionale
Le débit sanguin est fortement diminué dans
les territoires non concernés par l'exercice, et notamment le débit
splanchnique au profit des muscles en action qui peuvent ainsi décupler leur
débit de repos. Cette fonction permet d'alimenter le muscle en oxygène et de
drainer les déchets de son travail. Cette redistribution se fait après 3 à 4 mn
d'effort expliquant la nécessité d'un échauffement, pour alimenter les muscles
jusqu'à 15 fois le débit initial.

¥
De 15% au repos, le débit sanguin des muscles actifs passe à 80-85 % du débit
cardiaque.
¥
Le flux sanguin cérébral augmente au mieux, légèrement à l'effort.
¥
L'exercice entraîne une fuite liquidienne vers les muscles squelettiques en
activité et une réabsorption des liquides tissulaires en compensation. Durant
un exercice relativement intense sur bicyclette, 1100 ml fuient vers les
muscles, mais 500 ml sont repris aux tissus au repos.
III/ Adaptation de la distribution
sanguine locale
Pour pouvoir accepter l'augmentation du
débit sanguin musculaire et ravitailler toutes les cellules du muscle,
l'organisme ouvre les sphincters précapillaires dont seulement 1 sur 30 à 40
est ouvert au repos. L’ouverture de capillaires dormants au cours d’un exercice
sert trois buts:
¥ augmenter substantiellement le débit sanguin
intramusculaire
¥ augmenter le débit du fait de l’ouverture de
plusieurs vaisseaux, sans que la vitesse d’écoulement n’augmente beaucoup
¥ améliorer les échanges entre le sang et les cellules
musculaires par une meilleure irrigation
Cette vasodilatation est produite par
différents facteurs. La vasodilatation locale est directement proportionnelle à
l'intensité de l'effort. Le flux sanguin cutané diminue dans les cinq premières
minutes, puis, pour assurer l'élimination de la chaleur, augmente tant que la
charge de travail ne dépasse pas 50-60% de VO2 Max; il diminue
ensuite pour des efforts supérieurs au profit de la perfusion musculaire. En
effet, au delà de 50-60% de VO2 Max, le flux sanguin diminue
jusqu'au niveau maximal où il y a vasoconstriction, même en présence d'un
environnement chaud. Cette action permet au muscle au travail de pouvoir
multiplier par 50 son métabolisme. Les capillaires multiplient par 100 la
surface de diffusion en se dilatant à l'effort. Au cours d’un exercice
vigoureux, il peut y avoir plus de 4 000 capillaires ouverts par millimètre
carré de section musculaire transversale dans le muscle sollicité. Il a été
observé que le nombre de capillaires par muscle et le nombre de capillaires par
millimètre carré de tissu musculaire était de 40% plus élevé chez des athlètes
d'endurance que chez des sujets témoins non entraînés. Plus le nombre de
capillaires augmente, meilleure est l’oxygénation des fibres musculaires,
particulièrement dans le cas d’exercices vigoureux nécessitant beaucoup
d’énergie en aérobiose.
L'adaptation locale est quelque peu
différente pour les exercices de force. Lorsqu’un muscle se contracte à 60% de
sa capacité maximale, l’importante pression intramusculaire interrompt le débit
sanguin. C’est ce qui se produit au cours d’une contraction isométrique.
L’énergie nécessaire à la poursuite de l’effort est donc prélevée sur les
réserves de phosphagènes et de glycogène par des réactions anaérobies.
En fait, le degré de vasodilatation
intramusculaire est proportionnel à la force de contraction. Aux facteurs
locaux responsables de la vasomotricité s’ajoute un contrôle central
sympathique et, à un moindre degré, parasympathique. Par exemple, les muscles
renferment de petites fibres nerveuses sensibles aux substances libérées
localement au cours d’un effort. Sur stimulation de ces fibres, le système
nerveux central reçoit le message d’ajuster en conséquence le système
cardio-vasculaire. Les fibres sympathiques qui libèrent une substance
vasoconstrictrice sont dites adrénergiques. Les autres fibres sympathiques, et
spécialement celles qui innervent les tissus musculaire et cardiaque, libèrent
de l'acétylcholine; ce sont des fibres cholinergiques, vasodilatatrices.
L'augmentation du retour veineux
Différents mécanismes contribuent à
augmenter le retour veineux au cours des exercices: la pompe musculaire, la
pompe respiratoire et la veinoconstriction.
Ø
La pompe musculaire: durant une épreuve d’haltérophilie ou au cours
d’autres exercices isométriques, la pompe musculaire ne fonctionne pas et le
retour du sang veineux est plus difficile.
Ø
La pompe respiratoire: plus le rythme et le volume de la respiration
sont grands pendant un exercice, plus cette pompe est efficace.
Ø
La veinoconstriction: C‘est l’un des différents réflexes mis en jeu et
contrôlés par le système nerveux au cours des exercices. Au repos, seulement
entre 15 et 20% de tout le débit systémique est distribué aux muscles; lors de
l’exercice maximal, les muscles qui travaillent peuvent recevoir entre 85 et
90% du débit sanguin total.
IV/ Adaptation orthosympathique et
endocrinienne
 L'activité sympathique est augmentée lors de l'exercice. Un travail à 30%
de VO2 Max suffit à activer le sympathique et à libérer d'abord
noradrénaline et ensuite adrénaline entraînant une vasodilatation locale, une
accélération du rythme et de la puissance cardiaque, une augmentation du retour
veineux par vasoconstriction des veines assurant ce retour, une baisse des
résistances vasculaires systémiques (RVS).
L'activité sympathique est augmentée lors de l'exercice. Un travail à 30%
de VO2 Max suffit à activer le sympathique et à libérer d'abord
noradrénaline et ensuite adrénaline entraînant une vasodilatation locale, une
accélération du rythme et de la puissance cardiaque, une augmentation du retour
veineux par vasoconstriction des veines assurant ce retour, une baisse des
résistances vasculaires systémiques (RVS).
 Une augmentation du tonus vagal accroît le débit coronaire de 40%,
tandis qu'une stimulation du ganglion stellaire le réduit initialement pour
l'augmenter dans un second temps par l'intermédiaire d'une vasodilatation
coronaire.
Une augmentation du tonus vagal accroît le débit coronaire de 40%,
tandis qu'une stimulation du ganglion stellaire le réduit initialement pour
l'augmenter dans un second temps par l'intermédiaire d'une vasodilatation
coronaire.
L'inhibition sympathique détermine une
inhibition vasomotrice et une vasodilatation.
Il existe des fibres sympathiques
vasoconstrictrices adrénergiques qui constrictent les vaisseaux des organes abdominaux,
de la peau et des fibres sympathiques vasodilatatrices cholinergiques qui
dilatent dans le même temps les vaisseaux résistifs précapillaires des muscles
squelettiques. L'inhibition vasoconstrictive peut s'effectuer au niveau central
ou spinal.
L'action du système sympathique multiplie le
retour veineux par deux; associé à la vasodilatation locale musculaire le
retour veineux est multiplié par 5 ou 6.
L'effort intense, par le système
sympathique, augmente la production de granulocytes (système de défense non
spécifique).
Noradrénaline évolue jusqu'à 75% de PMA
Adrénaline évolue de 50% jusqu'à PMA
Le cortisol qui favorise le stockage du
glycogène augmente proportionnellement au-dessus de 60% de PMA
La production d’adrénaline et noradrénaline continue
à augmenter pendant les 3 mn suivant la fin de l’exercice. Cela peut expliquer
en partie les troubles du rythme post-effort.
Le cholestérol est à l’origine des hormones
stéroïdes
Des cortico-surrénales proviennent
l’aldostérone et le cortisol
Des gonades proviennent testostérone,
oestrogènes, progestérone.



V/ Adaptation de la tension
artérielle

 L'effort engendre par augmentation de l'activité orthosympathique:
L'effort engendre par augmentation de l'activité orthosympathique:
Ø une vasoconstriction veineuse qui améliore le retour
veineux
Ø une vasoconstriction artériolaire qui assure la
redistribution du débit cardiaque aux muscles actifs
Dans les territoires actifs, l'effet
vasoconstricteur est supplanté par l'effet vasodilatateur des métabolites
(adénosine) libérés localement induisant une chute des résistances vasculaires
systémiques de 266% chez un sujet normal, et une augmentation de la pression
artérielle moyenne du fait de l'augmentation du débit cardiaque. Il s'instaure
ainsi un gradient de pression entre le système artériel et le système veineux.
Chez le sujet sain, normotendu, au cours
d'un effort dynamique impliquant de grands groupes musculaires, la pression
artérielle systolique PAS augmente linéairement avec la charge et la
consommation d'O2 jusqu'à 180 mmHg à l'âge de 20 ans et 200 mmHg à
l'âge de 40 ans pour un effort modéré. Elle peut atteindre 250 Torrs, soit 250
mmHg. Cette augmentation de PAS est d'autant plus modérée que le sujet est
entraîné. L'augmentation de la tension, une minute après la fin de l'exercice,
signifie que le sujet a été au-delà du seuil anaérobie, et que l'épreuve était
presque maximale. Une diminution de l'activité sympathique après l'exercice
entraîne une diminution des résistances périphériques et explique la baisse de
PA.
 L'augmentation de PAS à l'exercice est la même pour les hypertendus que
pour les normotendus, cependant la pression artérielle est moins soumise aux
évolutions du rythme cardiaque pour des hypertendus s'entraînant en aérobie.
L'augmentation de PAS à l'exercice est la même pour les hypertendus que
pour les normotendus, cependant la pression artérielle est moins soumise aux
évolutions du rythme cardiaque pour des hypertendus s'entraînant en aérobie.
Une bonne contractilité explique des mesures
élevées de tension à l'exercice, et impliquent une bonne perfusion cardiaque.
La TA diastolique s'élève peu (10 à 15 Torrs
au maximum). TAD peut même diminuer légèrement à l'effort.
Nota: une étude récente donne une TAS
moyenne à l'effort de 160 à 170 mmHg. Une autre étude publiée donne des
résultats à 150 W entre
177 et 209 Torr chez les hommes et de 152 à
188 Torr chez les femmes. Les statistiques semblent néanmoins prouver que le
taux de mortalité est le plus faible quand TAS à l'effort égale ou dépasse 200
mmHg sans atteindre la limite de 250 mmHg pour un effort maximal; cependant des
augmentations de TAS au-delà de 220 mmHg doivent être considérées et vérifiées.
L’impossibilité pour la pression sanguine
d’augmenter à l‘effort peut refléter une dysfonction cardio-vasculaire. Ainsi,
l’impossibilité pour la pression systolique d’augmenter d’au moins 2,5 à 4 kPa
(20 à 30 mmHg) au cours d’un effort progressif peut refléter une diminution de
la réserve cardiaque. Une augmentation importante et rapide de la fréquence
cardiaque (tachycardie) au début de l’effort constitue souvent un signe
avant-coureur de problèmes cardiaques.
|
Evaluation des chiffres tensionnels à l'effort en fonction de l'âge et de la FC chez 190 sujets féminins normotendus |
||||||||||||||
|
Niveau travail |
20 - 29 ans |
30 - 39 ans |
40 - 49 ans |
50 - 59 ans |
||||||||||
|
|
FC |
TAS |
TAD |
FC |
TAS |
TAD |
FC |
TAS |
TAD |
FC |
TAS |
TAD |
||
|
repos |
83± 7 |
121± 8 |
79± 6 |
83± 7 |
123± 7 |
82± 7 |
83± 8 |
126± 7 |
82± 7 |
72± 9 |
123± 8 |
81± 8 |
||
|
25 W |
124± 6 |
146± 7 |
86± 5 |
118± 8 |
145± 7 |
87± 8 |
120± 8 |
149± 10 |
94± 9 |
107± 11 |
155± 10 |
91± 10 |
||
|
50 W |
135± 7 |
151± 8 |
86± 6 |
135± 8 |
157± 9 |
92± 10 |
129± 7 |
163± 10 |
97± 8 |
115± 9 |
166± 10 |
91± 11 |
||
|
75 W |
148± 9 |
162± 10 |
89± 7 |
152± 10 |
172± 10 |
95± 9 |
142± 9 |
178± 10 |
99± 9 |
137± 11 |
176± 12 |
95± 10 |
||
|
Evaluation des chiffres tensionnels à l'effort en fonction de l'âge et de la FC chez 198 sujets masculins normotendus |
||||||||||||||
|
Niveau travail |
20 - 29 ans |
30 - 39 ans |
40 - 49 ans |
50 - 59 ans |
||||||||||
|
|
FC |
TAS |
TAD |
FC |
TAS |
TAD |
FC |
TAS |
TAD |
FC |
TAS |
TAD |
||
|
repos |
79± 6 |
124± 6 |
80± 4 |
76± 7 |
128± 4 |
81± 3 |
74± 5 |
126± 5 |
82± 4 |
74± 6 |
130± 5 |
85± 3 |
||
|
50W |
106± 6 |
142± 7 |
82± 4 |
104± 7 |
147± 7 |
85± 4 |
100± 5 |
152± 7 |
87± 5 |
94± 9 |
158± 8 |
95± 6 |
||
|
75 W |
121± 8 |
152± 8 |
84± 3 |
116± 8 |
157± 6 |
87± 4 |
118± 8 |
172± 10 |
97± 5 |
109± 6 |
183± 8 |
92± 4 |
||
|
100 W |
135± 9 |
165± 10 |
85± 5 |
129± 7 |
173± 8 |
91± 10 |
123± 8 |
181± 13 |
94± 6 |
118± 7 |
187± 10 |
99± 6 |
||
|
125 W |
153± 10 |
181± 10 |
91± 7 |
145± 11 |
190± 7 |
91± 5 |
144± 8 |
198± 10 |
97± 6 |
130± 7 |
202± 12 |
103± 6 |
||
|
150 W |
157± 8 |
191± 14 |
91± 8 |
148± 10 |
197± 12 |
91± 4 |
147± 8 |
197± 8 |
95± 5 |
136± 7 |
199± 6 |
100± 6 |
||
La mesure se prend à l'état d'équilibre
(steady state), c'est-à-dire quand Fc est stable soit une variation £ 5
pulsations en 30 secondes.
Conditions tensionnelles d'arrêt de l'exercice
Ø
chute ou défaut d'augmentation de la tension artérielle systolique
Ø en-dessous de 5 cmHg pour une Fc Max ³ 150 bat/min
Ø en-dessous de 3 cmHg pour une Fc Max < 115
bat/min
Ø
augmentation anormale de la tension systolique
Ø ³ 22 cmHg pour 50% RFc
Ø ³ 25 cmHg pour un effort maximal
Ø ³ 28 cmHg pour un hypertendu actif en effort maximal
Ø
augmentation de la tension diastolique plus importante que l'augmentation de la
tension systolique
Ø
augmentation de la tension diastolique au delà de 1 cmHg
Ø
augmentation de PAS à 230 mmHg et PAD à 120 mmHg chez l'enfant
Nota: quand le patient atteint VO2
Max, TAS diminue reflétant la baisse de VES et du débit cardiaque, accompagnant
le début du travail en anaérobie.
Etudes
réalisées par Astrand sur l'évolution de la tension suivant le type d'effort
Comparaison des
pressions de 7 sujets pendant un travail
|
|
Contractions isométriques (% contraction isométrique maximale volontaire CMV) |
Développé charges inertes (% CMV) |
Développé résistance hydraulique |
|||||
|
Mesure |
25% |
50% |
75% |
100% |
25% |
50% |
Lent |
Rapide |
|
PAS (kPa) |
22.9 |
23.8 |
26.6 |
29.9 |
22.5 |
30.9 |
31.5 |
32.6 |
|
PAD (kPa) |
14.1 |
15.4 |
18.0 |
20.7 |
13.8 |
20.5 |
13.4 |
21.3 |
Comparaison des pressions
au cours d'un exercice des membres inférieurs ou supérieurs
|
|
PAS (kPa) |
PAD (kPa) |
||
|
% VO2 Max |
Membre supérieur |
Membre inférieur |
Membre supérieur |
Membre inférieur |
|
25 |
20.0 |
17.6 |
12.0 |
9.3 |
|
40 |
21.9 |
18.4 |
12.4 |
9.4 |
|
50 |
23.3 |
19.2 |
12.8 |
9.7 |
|
75 |
27.3 |
21.3 |
13.7 |
10.0 |
Modifications tensionnelles lors de la récupération
TAS diminue la 1ère minute, puis
remonte et diminue de nouveau progressivement jusqu'au retour à l'état initial
vers la 6ème minute.
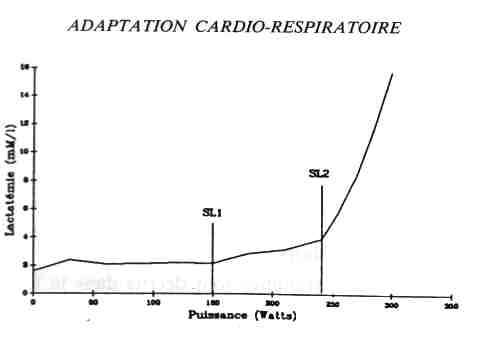

Au repos, la respiration nécessite 1% de la
consommation d'oxygène, à l'effort 12% de la consommation d'O2. Le volume
d'air respiré ou débit ventilatoire dépend de la fréquence respiratoire et du
volume courant. Au repos, il est de 5 à 7 litres par minute et s'accroît
proportionnellement à l'effort. Au cours d’un exercice maximal, l’augmentation
de la fréquence et de l’amplitude respiratoires peuvent entraîner des débits
aussi élevés que 200 litres par minute. Le volume courant dépasse rarement 55%
de la capacité vitale.
La ventilation maximale par minute de 160
l/min (homme) ou 110 l/min (femme) peut atteindre 400 l/min chez des sujets
entraînés. A l'effort maximum le travail s'effectue à 65-75% de la ventilation
maximale minute.
 Au repos, la fréquence respiratoire est comprise entre 10 et 20. Lors de
l'exercice, l'adaptation se produit d'abord par une augmentation du volume
courant qui peut atteindre 50% de la capacité vitale pour un travail d'une
intensité modérée. L'intensité de l'exercice entraînera ensuite une
augmentation de la fréquence respiratoire jusqu'à 40 voire jusqu'à 60 /min pour
des sportifs très entraînés. La fréquence respiratoire augmente linéairement
jusqu'à 80% de la capacité vitale et devient plus rapide à partir de ce point;
elle signe le seuil anaérobique traduisant la difficulté à éliminer l'acide
lactique. La fréquence respiratoire et le volume courant sont adaptés
spontanément par l'organisme pour le meilleur rapport efficacité / économie. Il
est préférable de respirer plus profondément que plus vite.
Au repos, la fréquence respiratoire est comprise entre 10 et 20. Lors de
l'exercice, l'adaptation se produit d'abord par une augmentation du volume
courant qui peut atteindre 50% de la capacité vitale pour un travail d'une
intensité modérée. L'intensité de l'exercice entraînera ensuite une
augmentation de la fréquence respiratoire jusqu'à 40 voire jusqu'à 60 /min pour
des sportifs très entraînés. La fréquence respiratoire augmente linéairement
jusqu'à 80% de la capacité vitale et devient plus rapide à partir de ce point;
elle signe le seuil anaérobique traduisant la difficulté à éliminer l'acide
lactique. La fréquence respiratoire et le volume courant sont adaptés
spontanément par l'organisme pour le meilleur rapport efficacité / économie. Il
est préférable de respirer plus profondément que plus vite.
Le contrôle de la respiration au cours de
l’effort ne dépend pas d’un seul facteur mais bien de la combinaison
probablement simultanée de plusieurs facteurs chimiques et nerveux (activation
corticale, influx des mécano-récepteurs articulaires et musculaires,
augmentation de la température).
On peut mentionner aussi un autre mécanisme
de contrôle de la ventilation. L’inflation des poumons durant l’inspiration
entraîne une expiration alors que la déflation des poumons durant l’expiration
stimule l’inspiration. Ces réflexes déclenchés par les récepteurs pulmonaires
sensibles à l’étirement assurent automatiquement le maintien de la fréquence
respiratoire.
Le point de côté serait en rapport avec une
ischémie diaphragmatique ou/et la présence de gaz intestinaux.
Le second souffle (disparition de la
difficulté à l'effort) est le stade d'adaptation ventilatoire à l'exercice et
celui de l'adaptation cardio-vasculaire des résistances périphériques; il
diminue les besoins en O2 du myocarde par diminution de la charge de travail.
Il faut de 10 mn entre 20 et 30 ans à 20-25 mn après 50 ans.
Après la fin de l’exercice musculaire il n’y
a pas de chute brusque immédiate du débit ventilatoire mais un retour très
progressif qui dure 5 minutes.
La production de dioxyde de carbone
Elle dépend de trois mécanismes:
Ø la production métabolique ou décarboxylation aérobie
résultant de la dégradation des substrats énergétiques au niveau du cycle de
Krebs. Au repos ou à l'effort de moyenne intensité en état stable, le
métabolisme oxydatif est mixte (glucides, lipides, protéines) et la production
de CO2 est inférieure à la consommation d'O2
Ø la production de dioxyde de carbone par la mise en
jeu du tamponnage par le bicarbonate. Lorsque l'intensité de l'effort entraîne
la participation de la filière aérobie lactique, le produit final de la
dégradation du glycogène en ATP est l'acide lactique, via le pyruvate. Cet
acide est décomposé en lactates et protons H+. Pour éviter une baisse du pH,
l'ion bicarbonate HCO3 tamponne le proton H+ pour former de l'acide carbonique
H2CO3 qui est lui-même dissocié par l'anhydrase
carbonique présente dans la cellule musculaire et dans le globule rouge en une
molécule d'eau H2O et une molécule de gaz carbonique CO2.
Ø L'hyperventilation est limitée généralement aux
premières minutes d'exercice (émotion, stress), et aux dernières (sprint).
 Quotient respiratoire = débit de CO2 / consommation d'oxygène
Quotient respiratoire = débit de CO2 / consommation d'oxygène
La dégradation aérobie des résidus glucosyls
entraîne un QR = 1.
La dégradation aérobie d'acides gras et d'acides
aminés entraîne un QR de 0,7 à 0,8.
Le métabolisme mixte du repos est entre 0,75
et 0,85 chez un individu normal.
Au début d'un effort, le QR va s'installer à
0,85 (hyperventilation de stress), puis descendre vers la valeur de repos dans
le cadre d'un exercice de moyenne intensité, avant de remonter vers l'unité
avec la consommation de glycogène à l'augmentation de l'effort (> 60% VO2
Max), signe d'une participation de la filière anaérobie lactique, pour ensuite
dépasser 1 au moment de l'hyperventilation traduisant la prépondérance des
filières anaérobie lactique et alactique (augmentation importante de l'effort
et sprint). Le QR peut atteindre 1,21± 12 à l'effort maximal.
On retrouve la notion de seuil anaérobique à
QR = 1, puisqu'en dessous l'oxygène combine avec l'acide lactique et au-dessus
l'acide lactique s'accumule faute de suffisamment d'oxygène pour l'excréter.
La pression partielle en O2 dans le
sang artériel se modifie peu (- de 10 Torrs) à l'effort chez un sujet sain et
reste supérieure à 80 Torrs (80 mmHg).
La pression partielle en CO2
comprise entre 35 et 45 Torrs varie peu à l'effort (± 2 Torrs),
jusqu'au seuil d'accumulation sanguine de lactates ou seuil anaérobique, où
elle peut chuter jusqu'à 25 Torrs pour 75% de VO2 Max.
La lactatémie de repos est voisine de 1
mmole/litre. Elle augmente jusqu'à 2 mmoles/l. à 50% de VO2 Max, et
jusqu'à 4 mmoles/l. à 75% de V02 Max. Au delà de cette zone qui
représente en général le seuil anaérobique, la lactatémie augmente
exponentiellement jusqu'à atteindre 7 à 12 mmoles/l. pour un sujet non entraîné
et 20 mmoles/l. et plus chez des sportifs de haut niveau très bien entraînés.
L'augmentation de la lactatémie entraîne une acidose avec baisse concomitante
de la réserve alcaline (HCO3) et du pH. Ce dernier, en fin d'effort maximal
peut atteindre 7,2 et même 6,9 chez des sujets bien entraînés.
Il faut se rappeler que le lactate pour
moitié rejoint le flux sanguin et pour l'autre moitié est oxydé par les fibres
lentes du muscle.
Discussion à propos du seuil anaérobie
Le seuil anaérobie ou seuil d'accumulation
sanguine des lactates est situé pour certains à 2 mmoles/l pour d'autres à 4
mmoles/l. En fait cette notion de seuil est floue puisque il dépend pour
beaucoup de facteurs personnels comme l'entraînement.
 Chez un individu moyen, 2 mmoles/l représente le seuil où la production
de CO2 et la ventilation externe (VE) décrochent de la parallèle à
la courbe de consommation d'O2 qu'elles suivaient. Ce seuil est
situé généralement à environ 50% de VO2 Max, toujours pour un
individu moyen.
Chez un individu moyen, 2 mmoles/l représente le seuil où la production
de CO2 et la ventilation externe (VE) décrochent de la parallèle à
la courbe de consommation d'O2 qu'elles suivaient. Ce seuil est
situé généralement à environ 50% de VO2 Max, toujours pour un
individu moyen.
4 mmoles/l situé en général à 75% de VO2
Max, est le seuil où la courbe de VE décroche de la parallèle à la production
de CO2 qu'elle suivait jusque-là. Ce seuil représente en fait, celui
où les mécanismes anaérobies prennent le pas sur les mécanismes aérobies; il
s'agit donc du seuil à partir duquel l'individu va s'épuiser le plus vite.
Cette notion de seuil anaérobique est théorique,
car le degré d'entraînement des individus en change les valeurs de manière
importante. De plus ces décrochages sont observés dans le syndrome de McArdle,
où le sujet est dans l'impossibilité de produire de l'acide lactique, et dans
le cas où un sujet sain a augmenté au préalable à l'effort sa lactatémie.
 L'augmentation de lactates représente donc la production par le
mécanisme anaérobie lactique et les fibres rapides d'une quantité de lactates
supérieure à ce que peut utiliser l'oxygène extrait par le muscle, les fibres
lentes et la réserve alcaline de HCO3.
L'augmentation de lactates représente donc la production par le
mécanisme anaérobie lactique et les fibres rapides d'une quantité de lactates
supérieure à ce que peut utiliser l'oxygène extrait par le muscle, les fibres
lentes et la réserve alcaline de HCO3.
Evolution des paramètres respiratoires
ERO2 (équivalent respiratoire
pour l'O2) = VE / VO2 et est compris entre 20 et 26 chez l'adulte
sain au niveau de la mer. Il faut donc ventiler entre 20 à 26 litres d'air pour
consommer 1 litre d'O2. ERO2 monte au début de l'exercice
entre 30 et 40 (hyperventilation de stress), pour redescendre entre 20 et 26
pour un effort à 50-60% de VO2 Max et augmenter linéairement à
l'intensité de l'effort à partir de ce seuil qui est le seuil des 2 mmoles/l de
lactates jusqu'à une valeur comprise entre 30 et 40 à l'effort maximum. Le
niveau de ventilation pulmonaire mesuré sur les gaz expirés (VE) est à l'effort
bien plus dépendant de la nécessité d'éliminer du CO2 que d'apporter
de l'O2. Lors d'un effort, le maintien du pH dépend du maintien de
la pression partielle en CO2 dans le sang artériel, donc de
l'élimination pulmonaire du CO2 produit au niveau musculaire.
L'information est transmise par les chémorécepteurs carotidiens.
VE d'un sujet sain atteint 120
litres/minute. Au delà, toute augmentation de VO2 par augmentation
de VE ne servirait qu'à satisfaire les besoins supplémentaires des muscles
respiratoires, et ne serait pas utilisée en périphérie. Le coût respiratoire
est en moyenne de 4% au repos et pour un effort modéré. Il peut atteindre
presque 10% pour un effort très intense.
L'efficacité de VE dépend du volume qui
participe aux échanges gazeux par rapport à celui qui ventile l'espace mort.
Cet indice se traduit par le rapport volume d'espace mort / volume courant soit
Vd / Vt. Plus ce rapport est bas, plus VE est efficace et proche de la
ventilation alvéolaire.
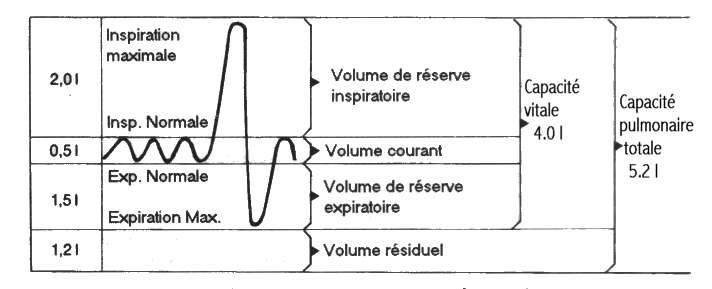
Chez un individu normal au repos, Vd / Vt
est compris entre 0,25 et 0,35. Il diminue à l'effort entre 0,20 et 0,05 parce
que le volume courant augmente proportionnellement plus que le volume d'espace
mort.
Chez un individu normal, l'équivalent
respiratoire pour le dioxyde de carbone ERCO2 ou rapport VE / VCO2
est de 22,1 litres. Il faut donc 22.1 l. supplémentaire de VE pour éliminer un
litre supplémentaire de CO2. Ce rapport est vérifié jusqu'à 75% de
VO2 Max (seuil anaérobique probable). Au delà le pH baisse et
stimule l'augmentation de VE, qui détermine une baisse de la pression partielle
artérielle en CO2 jusqu'à 25 Torrs à l'effort maximum. A ce niveau
très intense, la production de catécholamines, l'élévation de la température
interne, le stress de l'effort, contribuent à l'hyperventilation, où VE s'élève
plus rapidement que VCO2 permettant a ERCO2 d'atteindre à
l'effort maximum un rapport voisin de 32, soit 32 l. d'air pour ventiler 1 l.
de CO2.
Un sujet sain à l'effort maximum pourra
augmenter VE jusqu'à 70-75% de sa ventilation minute maximale (VMM).VMM = 35 x
VEMS (volume maximal expiré par seconde).
La réserve ventilatoire est la différence
entre la ventilation minute maximale et la ventilation expirée maximale mesurée
à l'effort. Elle est normalement supérieure à 15 litres par minute.

Le volume courant Vt d'un sujet normal est
proche de 6 ml/kg soit 450 ml pour un homme de 75 kg. Il comprend l'espace mort
anatomique Vd et le volume d'air arrivant aux alvéoles Va. On estime l'espace
mort à 2 ml/kg soit 150 ml pour le même homme de 75 Kg. A l'effort
l'augmentation de Vt est la première ressource d'augmentation de VE, la seconde
étant l'augmentation de la fréquence respiratoire fr. VE = fr x Vt.. Un
compromis s'instaure entre fr et Vt pour que la ventilation alvéolaire soit
assurée pour la dépense minimum d'O2. Ce compromis varie ensuite en
fonction des efforts et des groupes musculaires mis en action.
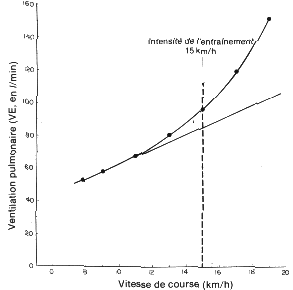 Vt peut augmenter jusqu'à 60 à 70% de la capacité vitale CV du sujet
sain.
Vt peut augmenter jusqu'à 60 à 70% de la capacité vitale CV du sujet
sain.
L'espace mort anatomique Vd s'accroît un peu
à l'effort jusqu'à 300-350 ml, mais Vd / Vt diminuera puisque Vt augmentera
proportionnellement plus que Vd.
Les sommets des poumons sont au repos mieux
ventilés que perfusés chez un sujet debout. L'augmentation du débit cardiaque à
l'effort augmente la pression dans l'artère pulmonaire et aboutit donc une
meilleure perfusion des zones sous-perfusées par recrutement des capillaires
précédemment collabés ou partiellement dilatés.
fr est comprise entre10 à 20 cycles par
minute. A l'effort elle augmente moins que Vt tant que l'effort est inférieur à
75% de VO2 Max. Lorsque Vt atteint les deux tiers de la capacité
vitale, l'augmentation de VE est alors assurée par celle de fr. fr de fin
d'effort chez le sujet sain est comprise entre 30 à 50 cycles par minute; elle
peut monter jusqu'à 100 cycles chez le décompensé respiratoire. Pour une VE
déterminée, plus fr est basse, meilleure est la ventilation alvéolaire.
Un sujet fournissant un travail intense
respire d'avantage par la bouche, puisque la résistance à l'écoulement des gaz
est moindre que par le nez. Il est préférable de respirer par le nez, car l'air
est préalablement filtré et réchauffé et de ne respirer par la bouche qu'au
moment où la sollicitation devient trop importante. A partir de 40 l/mn le
sujet est obligé de respirer par la bouche.
VII/ Adaptation de la consommation
d'oxygène
La consommation d'oxygène (250 ml d'O2
au repos en moyenne) augmente linéairement avec l'intensité de l'effort jusqu'à
un certain seuil où elle se stabilise malgré l'augmentation de l'effort (VO2
Max) traduisant la puissance aérobie de l'individu. Un sujet sain peut
multiplier le débit de repos par 10 alors qu'un sujet très entraîné peut le
multiplier par 20 et plus. Au delà, l'exercice peut être maintenu jusqu'à
épuisement complet entre 3 et 15 minutes. Cette durée représente l'endurance
anaérobie de l'individu qui peut-être augmentée par l'entraînement dans cette
zone.
VO2 est plus élevée de 30% dans
un exercice effectué avec les jambes par rapport aux bras, donc Fc est plus
élevée par un travail de bras que par un travail de jambes à intensités
d'effort égales.
Il est important de se rappeler qu‘on impose
une plus grande charge métabolique et physiologique quand le travail recommandé
est effectué avec les membres supérieurs.
VO2 est légèrement plus élevée
sur tapis roulant que sur cycloergomètre.
l'augmentation de VO2 dépend du
débit cardiaque (FC x VES) et de la différence artério-veineuse en O2
(capacité d'extraction de l'O2 du sang)

Le début de l'exercice est anaérobique et
l'adaptation ventilatoire se produit en 3 à 4 minutes. Ce retard dans la
consommation d'oxygène par rapport aux besoins théoriques qui se constituent au
début de l'exercice est restitué après arrêt de l'effort grâce à une
consommation supérieure aux besoins. Ce remboursement d'oxygène est toujours
supérieur à la dette d'oxygène, car il comprend l'oxygène nécessaire à certains
mécanismes (comme la retransformation de l'acide lactique).
Un délai de 3 minutes est nécessaire pour
atteindre la consommation maximale d’oxygène car le délai d’apparition de la
consommation maximale d’oxygène par l’organisme provient de l’inertie des
adaptations cardio-vasculaires et tissulaires à l’effort:
Ø augmentation du débit cardiaque,
Ø modification de la répartition de la masse sanguine
corporelle,
Ø augmentation du débit sanguin musculaire,
Ø activation des réactions d’oxydation de la fibre
musculaire.
 Si le temps de latence est de trois à quatre minutes chez le sédentaire,
il est proche de 90 secondes chez le sportif entraîné et échauffé.
Si le temps de latence est de trois à quatre minutes chez le sédentaire,
il est proche de 90 secondes chez le sportif entraîné et échauffé.
VIII/ Amélioration de la différence artério-veineuse
Le muscle au repos ne prélève qu'une partie
de l'oxygène et des substrats apportés par le sang artériel. A l'effort le
muscle augmente son extraction sanguine en oxygène et en substrats.

L'entraînement augmente la capacité
d'extraction de l'oxygène par l'organisme. De 4 à 5 ml O2 pour 100
ml de sang, la différence artério-veineuse à l'effort peut quadrupler chez un
sportif très entraîné. En effet, si l'extractiond'O2 est de 5% au
repos, elle est portée à l'effort maximum à 10-12% chez l'individu non entraîné
et à 16-18% chez l'individu entraîné.
Le pouls d'oxygène = VO2
/ Fc = Qc x (CaO2- CvO2) / Fc
Ce rapport indique la quantité d'O2
apportée à chaque battement cardiaque soit en moyenne 16 ml O2 / 100
ml sang au repos.
IX/ Adaptation de la courbe de
dissociation de l'oxyhémoglobine (HbO2)
La presque totalité de l'oxygène circulant
est liée à l'hémoglobine (Hb) sous forme d'oxyhémoglobine (HbO2). 1g
d'Hb peut se combiner à 1.3 ml d'O2. Le taux d'Hb dans le sang est
donc essentiel à l'oxygénation de l'organisme. L‘hémoglobine, protéine porteuse
de fer du globule rouge augmente d’environ 65 fois la capacité sanguine de
transport d’O2 par rapport à la quantité dissoute dans le plasma.
Chez l’homme, on retrouve environ 15 à 16
g d’hémoglobine par 100 ml de sang. Les femmes en possèdent légèrement
moins, 13 g d'Hb pour 100 ml de sang soit 4 g/100 ml de sang. Les femmes
qui ont un taux d'Hb moindre que celui de l'homme ont donc des capacités
physiques moindres. Pour cette raison, les compléments alimentaires en fer sont
très fréquents dans les milieux sportifs féminins.
Chaque gramme d’hémoglobine peut retenir
temporairement 1,34 ml d’oxygène. Quand l’hémoglobine est totalement saturée
d’oxygène, le sang peut transporter environ 20 ml d’oxygène par 100 ml.
Cependant, une diminution de la quantité de fer dans le globule rouge, comme
c'est le cas dans l’anémie ferriprive, réduit d’autant la capacité sanguine de
transport d’oxygène.
L'effet Bohr déplace vers la droite la
courbe de dissociation de l'oxyhémoglobine en réponse à toute augmentation de
l'acidité, de la température ou de la concentration de gaz carbonique.
La myoglobine est une protéine porteuse de
fer que l’on trouve dans les muscles squelettiques et le muscle cardiaque. Les
fibres musculaires rouges en possèdent beaucoup tandis que les fibres blanches
en sont dépourvues. La myoglobine ressemble à l’hémoglobine en ce qu’elle
s’associe de façon réversible à l’oxygène; toutefois, chaque molécule de
myoglobine ne contient qu’un seul atome de fer, et non quatre comme la molécule
d’hémoglobine. La myoglobine est une source supplémentaire d’oxygène pour le
muscle.
Les réserves d'oxygène liées à la myoglobine
fournissent 20% de l'énergie dépensée pendant une heure d'exercice intermittent
comportant un nombre égal de périodes de travail et de récupération de 15
secondes chacune.
Au repos, il n’y a qu’environ 25% de tout
l’oxygène qui soit libéré pour les tissus; le reste (75%) retourne au cœur par
les veines.
Un sujet normal au repos tient 14.9 g Hb
/100 ml sang et une saturation de 97,5% soit 20 ml d'O2 pour 100 ml
de sang. Une faible quantité d'O2 dissous (0.3%) est également
présente dans le plasma. Le contenu veineux en oxygène chez un sujet normal au
repos sera voisin de 14,62 ml O2/100 ml pour une saturation normale
de 72.5% et une quantité en O2 dissoute réduite.
D'une valeur de repos de 25% du contenu
artériel en O2, l'extraction d'oxygène passe à 70-80% du contenu
artériel en O2 à l'effort. A l'effort maximum, le contenu artériel
en O2 peut augmenter de 10% sous l'effet de l'hémoconcentration. A
l'effort chez le sportif entraîné le contenu veineux peut diminuer jusqu'à 2 ml
O2/100ml. La différence artério-veineuse maximum est atteinte pour
70-80% Fc Max.
A l'effort, l'élévation de la température au
delà de 40°C (au niveau du muscle actif), l'augmentation de la pression CO2
et la baisse du pH, influencent la différence artério-veineuse.
Le dopage sanguin par prélèvement de 3 à 4
unités de sang (450 ml chaque) en 3 à 8 semaines, extraction du plasma et
réinjection permet d'augmenter le nombre de globules rouges et donc l'oxygène
transporté. Les globules prélevés sont réinjectés 8 jours avant une épreuve
d’endurance. L'augmentation de l'hémoglobine de 8 à 20% se maintient pendant 14
jours minimum.
X/ Les facteurs limitants de la
performance
Les facteurs les
plus limitants de la performance sont
Ø le débit cardiaque
Ø la capacité de transport de O2
Ø la capacité périphérique d'extraction de O2
Ø le besoin en O2 des muscles respiratoires
La perte
d’électrolytes est facteur d’arrêt de l’exercice
On pense actuellement que le glucose
plasmatique ne participe pas massivement au métabolisme énergétique des
cellules musculaires et que son principal rôle énergétique concerne le système
nerveux (60%). La cellule musculaire est imperméable au glucose plasmatique qui
ne pénètre que grâce à l’action facilitante de l’insuline; or le taux
plasmatique d’insuline diminue parallèlement à la glycémie au cours de l’effort
modéré prolongé. C’est un phénomène très heureux car le glucose plasmatique est
ainsi réservé aux neurones qui ne peuvent pas utiliser d’autres substrats
énergétiques. La diminution des réserves de glycogène joue un rôle limitant
dans l’effort prolongé de plus de 75 minutes. En pratique, la consommation de
glucose pendant l’effort aide à épargner le glycogène musculaire et à retarder
la baisse de sucre dans le sang.
L’acidose métabolique n’est pas le seul
facteur limitant l’effort prolongé. L’épuisement des réserves de glucose,
l’excès de chaleur stockée par l’organisme peuvent également limiter une
épreuve d’intensité inférieure à la puissance maximale aérobie.
Au cours d’un exercice submaximal prolongé,
la fatigue s’installe d'abord par diminution du glucose circulant destiné au
SNC. Le facteur fatigue est donc essentiellement du à une hypoglycémie
cérébrale. A l'appui de ce fait, les performances en endurance d'un chien
perfusé en glucose au niveau carotidien ont été très nettement améliorées. La
fatigue est ensuite due à la réduction substantielle du glycogène musculaire.
Cette fatigue par défaut de nutriments survient même en présence d‘un apport suffisant
d’oxygène pour produire de l’énergie en aérobiose.
Au cours d’un exercice maximal la fatigue
correspond à une diminution de l’activité nerveuse, comme le révèle
l’électromyogramme. Cette fatigue est causée partiellement par un défaut de
transmission nerveuse ou neuro-musculaire.
Au cours d’un exercice maximal de courte
durée, la fatigue musculaire est liée au manque d’oxygène et à l’augmentation
de la concentration plasmatique et musculaire d‘acide lactique et donc, de la
concentration d’ions hydrogène dans les muscles à l’effort. Cette condition
anaérobie peut engendrer de fortes modifications intracellulaires dans les
muscles sollicités, dont une interférence avec la mécanique contractile, une
déplétion des réserves de phosphates riches en énergie, une altération du
transfert énergétique dans la glycolyse par suite d’une réduction de l’activité
d’enzymes clés, un dérangement du système réticulaire responsable de la
transmission de l’influx dans toute la fibre musculaire et un déséquilibre
ionique. La fatigue peut aussi se manifester à la jonction neuro-musculaire,
quand celle-ci ne peut laisser passer l’influx des motoneurones. La fatigue
centrale engendrant une inhibition des structures nerveuses centrales
responsables de la motricité, de ce fait, l'exercice ne pourra être prolongé
qu'avec l'aide de la musculature de soutien et des muscles synergiques, ce qui
augmentera la dépense énergétique et augmentera le nombre d'erreurs possibles.
Le mécanisme de l’installation de cette " fatigue nerveuse" est inconnu.
L'alcool et la nicotine engendre également
ce type d'inhibitions (stimulation de la formation réticulée).
Au cours d’un exercice submaximal prolongé,
de nouvelles unités motrices entrent en action afin de maintenir le niveau de
tension requise pour l’activité en question.
Un individu peut rarement dépasser 750 Watts
de dépense physique pendant une journée de travail de 8 heures. Pour pouvoir
travailler 8 heures en continu, un individu ne doit pas utiliser plus:
Ø de 30 à 40 % de sa puissance aérobie maximale
Ø 40 % de sa force maximale.
La motivation est nécessaire dès que le
sujet puise dans les réserves ordinaires, soit dès 35% de la puissance
maximale. La motivation permet de poursuivre 15 mn et plus un travail à VO2
Max qui serait normalement arrêté au bout de 4 à 5 mn.
Les réserves autonomes protégées ne peuvent
être utilisées que dans des conditions particulières (hypnose, compétition,
instinct de survie, colère etc...), car elles déterminent un épuisement total
pouvant aboutir à la mort.
Des concentrations d’acide lactique aussi
élevées que 200 mg/ 100 ml ont été mesurées chez les athlètes après
compétition. La fatigue ressentie par ces coureurs à la fin de la course est,
par conséquent, due à des facteurs autres que l’accumulation de l'acide
lactique. Quelques-uns des facteurs associés à ce genre de fatigue sont :
l une glycémie (concentration de glucose dans le sang)
basse causée par la déplétion des réserves de glycogène du foie
l une fatigue locale due à la déplétion des réserves de
glycogène musculaire
l une perte d’eau (déshydratation) et d’électrolytes,
qui induisent une augmentation de la température corporelle
l l'ennui dû à la monotonie de l’exercice.
Le contenu en eau d’un individu varie de 40
à 60% de sa masse corporelle. Sa teneur est de 65 à 75% dans le tissu
musculaire mais moins de 25% dans le tissu adipeux.
62% de l’eau corporelle est intracellulaire.
Environ 2,5 l. d’eau sont normalement nécessaires chaque jour à un individu
relativement sédentaire dans un environnement normal. L’eau vient de trois
sources:
 Ø boissons: un individu moyen
boit habituellement 1200 ml d’eau chaque jour. Cette quantité peut être
multipliée par 5 ou 6 fois lors d’exercice physique ou de stress thermique;
Ø boissons: un individu moyen
boit habituellement 1200 ml d’eau chaque jour. Cette quantité peut être
multipliée par 5 ou 6 fois lors d’exercice physique ou de stress thermique;
Ø
aliments;
Ø
métabolisme: lorsque les molécules d’aliments sont dégradées pour fournir de
l’énergie, il y a formation d’eau et de gaz carbonique. Cette eau d’origine
métabolique ou endogène comble environ 25% des besoins quotidiens d’un individu
sédentaire. La dégradation complète de 100 g de glucides, de protides et de
lipides produit 55, 100 et 107 g d’eau respectivement. Nous avons vu que chaque
gramme de glycogène retenait 2,7 g d’eau.
La thermorégulation est effectuée par
l'hypothalamus. L'organisme perçoit principalement le chaud grâce à la
température du sang irriguant l’hypothalamus.
Les thermorécepteurs cutanés sensibles au
froid détournent la circulation en profondeur. La production de frissons permet
de dégager de la chaleur. La régulation se fait plus par chaleur emmagasinée
que par chaleur produite. L'exercice au froid est plus coûteux que l'exercice à
la chaleur. Le froid augmente la production d'épinéphrine, de norépinéphrine,
thyroxine qui élèvent le métabolisme de repos, d'adrénaline et noradrénaline.
La chaleur corporelle peut se perdre par
radiation, par conduction, par convection et par évaporation. L'évaporation
constitue le principal moyen de protection contre la surchauffe. 1 l. d'eau
évaporée = 2425 Kj (580 Kcal). 75% de la chaleur est évacuée par convection et
radiation et 25% par évaporation dont 2/5ème par l'expiration et 2/5ème
par la sudation. L'action thermique d'une fenêtre ou du ciel sur l'individu est
l'irradiation.
Quand il vente ou quand on nage, le fait de
remplacer l’environnement par un autre toujours plus froid augmente les pertes
de chaleur, comme le fait de nager dans une eau froide.
L'évaporation de la sueur est favorisée par
la mobilité de l'air sauf si cet air est à une température supérieure à celle
de la peau.
L'enveloppe convective d'air chaud qui
entoure le corps peut aller jusqu'à plus d'un mètre au dessus du corps. En
position couchée, le débit de chaleur par convection monte au dessus de tout le
corps et la perte de chaleur par convection est d'environ 30% supérieure à celle
en position debout.
On compte environ 3 millions de glandes
sudoripares, contrôlées par les fibres cholinergiques du système nerveux
sympathique, qui excrètent de grandes quantités d’une solution légèrement
saline (hypotonique de 0,2 à 0,4% NaCl). Quand la sueur se retrouve sur la
peau, son évaporation cause un refroidissement; la peau ainsi refroidie
contribue au refroidissement du sang dirigé vers la périphérie. En plus de la
chaleur perdue par évaporation de la sueur, on évalue à 350 ml la quantité d’eau
qui s’évapore par transpiration dans une journée et 300 ml d’eau s’évaporent
par les voies respiratoires. Le taux de sudation augmente proportionnellement
avec la température ambiante et l'intensité du travail.
La présence de liquide sur la peau ralentit
la sudation. Les substances laissées sur la peau par la sueur évaporée
augmentent la pression osmotique de la surface cutanée et augmentent le débit
sudoral.
La capacité de
sudation des individus âgés et des enfants est limitée.
L‘évaporation de la sueur sur la peau dépend
de trois facteurs:
Ø la surface cutanée exposée à l’air atmosphérique
Ø la température et l’humidité de l’air ambiant
Ø les courants d’air au contact de l’organisme
L‘humidité relative est de loin le facteur
le plus important. L'humidité limite l'évaporation de la sueur car l'air est
déjà saturé, donc l'organisme produit encore plus de sueur. Quand le stress
thermique est énorme, 15 à 25% du débit cardiaque passe par la peau; une
seconde et demie après le début d’un exercice vigoureux, la sudation a déjà
débuté.
La perte de chaleur due à l’évaporation se
fait seulement lorsque le vêtement est entièrement mouillé. S'asperger d'eau
est donc un excellent moyen de lutter contre l'échauffement. Un costume sec ne
fait que prolonger la période entre la sudation et le refroidissement. Le coton
et la laine absorbent vite l’eau. Les gilets de sudation sont une aberration,
car ils empêchent l’air d’évaporer l’eau et donc de refroidir le corps. La
sueur doit pouvoir s'évaporer pour provoquer le refroidissement du corps. Moins
le corps est couvert, meilleure est l'évaporation. En ambiance chaude et humide
dont la température est inférieure à celle de la peau, il est nécessaire d'être
très peu vêtu. A l'inverse quand la température ambiante est supérieure à celle
de la peau, il vaut mieux adopter des vêtements amples, laissant circuler
l'air. L'air est un très bon isolant et les vêtements retenant l'air sont
indiqués pour assurer l'équilibre thermique.
 Le meilleur vêtement serait celui qui en ambiance de température
inférieure à une valeur donnée laisserait la chaleur se dissiper, et dans une
ambiance de température supérieure à la valeur, opposerait son air à la chaleur
produite.
Le meilleur vêtement serait celui qui en ambiance de température
inférieure à une valeur donnée laisserait la chaleur se dissiper, et dans une
ambiance de température supérieure à la valeur, opposerait son air à la chaleur
produite.
Chez le sujet habillé, la surface des mains (5%
de la surface corporelle) assure de 10% jusqu'à 20% de la dissipation de la
chaleur produite. L‘application de froid rafraîchit le sang en périphérie, ce
qui facilite la déperdition de chaleur (compresses ou douches froides).
La chaleur et les pertes d'eau
pendant l'exercice
Le principal danger encouru pendant l'exercice intense est celui
résultant de la perte d'eau non compensée. Si 500 à 700 ml de sueur sont
excrétés quotidiennement de manière normale, cette quantité ne correspond
absolument pas à la capacité de sudation lors d’un exercice prolongé par temps
chaud (1 à 3 l/h). En une à deux heures d'exercice intense, l'organisme peut
perdre jusqu'à 7 litres d'eau (7 kg). Le défaut d'évaporation entraîne une
élévation rapide de la température du corps, et dès 40°C des dommages
irréversibles peuvent se produire. Le cœur travaille moins bien en ambiance
chaude et pire encore si elle est humide. Si la température corporelle dépasse
41.7°C, un coup de chaleur peut survenir sous l'influence de perturbations du
SNC. Pour 200 g de perte d'eau à l'effort, le renouvellement spontané est de
95%. Pour 500 g de 75%, pour 750 g de 55%
Le principal objectif du réapprovisionnement
en eau est de maintenir le volume plasmatique à son niveau, afin d’assurer le
meilleur fonctionnement de la circulation et de la sudation. L'athlète bien
hydraté s’exerce toujours à un plus haut niveau qu’un athlète déshydraté; il
serait donc sage de boire 400 à 600 ml d’eau froide, 10 à 20 minutes avant un
exercice en ambiance chaude. Dans certaines activités comme la course de fond,
il est pratiquement impossible d’équilibrer l’approvisionnement à la perte, car
l’estomac ne peut laisser passer qu’environ 800 mI d’eau par heure au cours
d’un exercice vigoureux alors que la perte peut égaler 2 l./h.
C‘est l’intensité relative de travail qui
détermine l'augmentation de la température centrale au cours d’un exercice. A
la température cutanée normale 580 Kcal sont nécessaires pour évaporer 1 litre
de sueur. Le rendement mécanique de l'exercice étant de 25%, 75% de l'énergie
est transformée en chaleur. Une production de 50 Kcal suffit à augmenter la
température du corps de 1°C en l'absence de système de refroidissement.
Une perte d'eau au delà de 3% du poids de
corps affecte significativement la performance. Si un sujet n'a pas compensé
80% de sa perte d'eau de la dernière séance ou de plus de 3% de son poids de
corps entre deux séances espacées au maximum de 48h, il faut surseoir à la
deuxième séance, car il existe un danger pour le sujet puisque le volume
sanguin est réduit quand la sueur produite atteint 2 à 3% de la masse
corporelle.
Une perte d'eau de 10% du poids de corps
peut affecter dangereusement la santé du sujet. L'échauffement excessif de
l'organisme provoque la mort.
Une déshydratation correspondant à 5% de la
masse corporelle entraîne une augmentation significative de la température
rectale et de la fréquence cardiaque, et une réduction marquée de la sudation
chez des hommes et des femmes qui travaillent dans une ambiance confortable ou
chaude. Physiologiquement, les adultes tolèrent assez bien un déficit hydrique
de 1,5 litre. Quand la perte d’eau correspond à 4-5 % de la masse corporelle
(phénomène fréquent chez les lutteurs de niveau secondaire), il y a une
réduction nette de la capacité de travail physique; les fonctions organiques
sont également altérées. Des sujets déshydratés jusqu'à 4,3% de leur masse
corporelle ont présenté 48% moins d’endurance à la marche; leur V02
Max était réduit de 22%. Dans cette même série d’expériences, on observa une
diminution d’endurance de 22% et une diminution de VO2 Max de 10%
quand le niveau de déshydratation ne représentait que 1 ,9% de la masse
corporelle.
La perte d'eau dépend de la température et
de l'humidité ambiante. Plus l'atmosphère est humide, moins la sueur peut
s'évaporer. Une humidité relative de 95% est dangereuse, car il devient très
difficile d'éliminer la chaleur par évaporation. A 100% d'humidité relative,
l’air est totalement saturé d’eau. L’évaporation de la sueur sur la peau est
donc impossible.
Le métabolisme de repos observé sous un
climat tropical est habituellement 5 à 20
fois plus élevé que celui retrouvé en milieu
tempéré, car chaque fois que la température s'élève de 1°C l'intensité des
métabolismes s'accroît de 13%. L’exercice en ambiance chaude stimule le
métabolisme; le même exercice accompli en ambiance thermiquement neutre
entraîne une diminution d’environ 5% de la consommation d’oxygène, probablement
due à une plus haute température interne et à la plus grande quantité d’énergie
requise pour le fonctionnement des glandes sudoripares et aux ajustements
circulatoires du travail en ambiance chaude.
 La température ambiante idéale d'un individu au repos est de 20°; une
température entre 21 et 24° représente le confort optimal, pour une température
cutanée optimale de 33°C. Cette température doit baisser avec l'intensité de
l'exercice pour aboutir à 15° en cas d'exercice intense.
La température ambiante idéale d'un individu au repos est de 20°; une
température entre 21 et 24° représente le confort optimal, pour une température
cutanée optimale de 33°C. Cette température doit baisser avec l'intensité de
l'exercice pour aboutir à 15° en cas d'exercice intense.
Le travail dans un environnement chaud
augmente les lactates, car une partie du sang destiné aux muscles est détourné
vers la peau pour assurer le refroidissement. L'exercice par temps chaud
sollicite plus l’anaérobie que par temps froid.
L‘eau perdue par l’usage d’un diurétique
provient davantage du plasma que des autres compartiments liquidiens. Les
pertes de potassium peuvent être considérables et affaiblir les muscles. Les
diurétiques peuvent grandement affecter les fonctions neuro-musculaires. Suer
pour maigrir est une hérésie dangereuse, hérésie parce que la graisse ne fond
pas, et l'eau perdue se reconstitue, dangereuse du fait de la déshydratation.
Il n’est pas établi qu’un apport
d'électrolytes, au cours d‘un exercice par temps chaud, améliore la performance
ou réduise le stress physiologique, dont les crampes musculaires.
La sueur est hypotonique et la sudation
détermine une perte d'eau supérieure à la perte de sel. L'ingestion de NaCl est
donc contre-indiquée sauf en cas de sudation importante associée à une activité
prolongée. L'organisme est capable de réguler son excrétion en Na. Il n’y a pas
de raison de consommer des boissons dites "athlétiques" pour
remplacer les minéraux perdus par sudation. Quand un individu fait beaucoup
d’exercices par temps chaud, la sueur peut entraîner 13 à 17 g de sel (2,3 à
3,4 g/l de sueur) quotidiennement. C’est environ 8 g de plus de ce que fournit
l’alimentation. Dans ce cas, il faudrait un supplément de sel de 0,1 à 0,2%,
soit environ un tiers de cuillère à thé de sel de table dans un litre d’eau. Il
n’est pas certain qu’il faille ajouter du potassium, car les pertes dues à la
sudation sont négligeables, à moins de conditions extrêmes. Un verre de jus
d’orange redonne à peu près tout le potassium, le calcium et le magnésium
emportés par 2 à 3 litres de sueur. C'est par des modifications diététiques et
le travail des reins que l’organisme compense la perte d’électrolytes par la
sudation. En fait, les études scientifiques démontrent que les individus
ajustent inconsciemment leur apport de sel quand le besoin s’en fait sentir.
Le chrome et le zinc, oligo-éléments
nécessaires au bon fonctionnement des métabolismes glucidique et lipidique,
peuvent être chassés de l’organisme à la suite d’un exercice vigoureux.
Comme la sueur excrétée entraîne de l’eau et
des électrolytes, l’organisme faisant face à un stress thermique met en jeu des
ajustements hormonaux pour conserver le plus d’eau et de sels possible.
L’hypophyse gère l’hormone antidiurétique (ADH) pour augmenter l'absorption
rénale d’eau. C’est pourquoi l’urine est plus concentrée au cours d’un stress
thermique. Après plusieurs jours d'un exercice en ambiance chaude, ou au cours
d’une seule séance d’exercice, le cortex surrénal libère l’aldostérone, une
hormone qui préserve le sodium. La concentration de sodium dans la sueur
diminue afin de ménager les électrolytes à mesure que l’organisme est exposé à
un stress thermique.
Une perte excessive de liquide et de sels
minéraux peut aussi survenir en ambiance froide.
Prévention des troubles liés à la
chaleur
Ø
L’athlète devrait noter sa masse le matin (immédiatement après le lever du lit,
et après avoir uriné, mais avant le petit déjeuner) pour détecter les symptômes
d’une déshydratation chronique. Le seul moyen réel de quantifier les pertes
d'eau et les quantités à reconstituer est la pesée avant et après exercice.
Ø
S'assurer grâce à un psychomètre de la température ambiante et du degré
d'humidité, et éventuellement reporter l'entraînement si nécessaire:
Ø Moins de 15°C: aucune précaution à prendre;
Ø 15°à18°C: surveiller attentivement tous les joueurs,
surtout ceux qui perdent beaucoup de poids;
Ø 18° à 21°C: insister pour que les joueurs boivent;
Ø 21°à 24°C: modifier la séance pour accorder des
périodes de repos toutes les 30 minutes, en plus d’observer les précautions
énumérées précédemment;
Ø 24°C ou plus: reporter l’entraînement ou y prendre
part en tenue légère.
Ø Les courses de distance (16 kilomètres et plus) ne
doivent pas avoir lieu lorsque la température mesurée au thermomètre humide
excède 28°C.
Ø
La soif ne suffit pas à indiquer les dépenses d'eau à reconstituer. La
sensation de soif intervient en retard, et il faut anticiper pendant l'exercice
les pertes d'eau. la sensation de soif débute après une perte de 0.5 à 1 l.
Ø
Boire 400 à 600 ml d’eau ou d’une boisson spécifique (sans glucides rapides),
30 minutes avant le départ de la compétition par dose maximum de 250 ml par ¼
h.
Ø
Pendant la compétition, ingérer par ¼ h un maximum de 250 ml d'eau ou d’une boisson
spécifique (légèrement glucosée admis). L'organisme met en effet 15 minutes à
assimiler 250 ml d'eau. Au delà cette eau reste dans l'estomac et gêne le
sportif pendant son exercice.
 Ø La rapidité d'ingestion
d'une solution glucosée n'est pas gênée par l'exercice si son intensité est
inférieure à 70% deVO2 Max
Ø La rapidité d'ingestion
d'une solution glucosée n'est pas gênée par l'exercice si son intensité est
inférieure à 70% deVO2 Max
Ø
Inciter les sportifs à boire pendant l'exercice surtout ceux qui se
déshydratent le plus. Il faut que les sportifs puissent avoir facilement de
l'eau à leur disposition. Les organisateurs de la course devraient installer
des postes d’eau tous les 3 à 4 km pour les courses de 16 km et plus; la
boisson joue un rôle significatif dans les courses prolongées de plus de 50 à
60 minutes.
Ø
Après la compétition, un régime sodé modéré et l’ingestion de boissons
contenant les sels minéraux essentiels peuvent remplacer adéquatement les
électrolytes (sodium et potassium) perdus dans la sueur.
.
Lorsque deux boissons ont la même pression
osmotique, elles sont dites isotoniques. Quand une boisson a une pression
osmotique supérieure à une autre, elle est dite hypertonique. Or deux liquides
séparés par une membrane possédant deux pressions osmotiques différentes
tendent à équilibrer cette pression par passage de la solution hypotonique vers
la solution hypertonique.
En conséquence, une boisson glucosée étant
un liquide hypertonique par rapport au plasma sanguin, c'est l'eau du plasma
qui est attiré vers l'estomac pour diluer la solution glucosée. Il faut donc
déterminer, après un exercice, quel est l'élément reconstituant le plus
important de l'eau ou du sucre et privilégier des solutions non glucosées (eau
pure) en cas de déshydratation ou faiblement glucosées (< 60 g/l) en cas
d'hypoglycémie.
Concentration
de glucose selon la température ambiante:
< 5°C 80 à 100 g/l
de 5 à 15°C 65 g/l
de 15 à 25°C 40 g/l
> 25°C au maximum 30 g/l
L'eau ingérée doit être 25 cl (12 ml/kg/h)
d'une solution glucosée à 25 g/l (½ sucrose, ½ fructose) d'une température de
10 à 15°C toutes les quinze minutes et de moins de l0 mEq/l ou 230 mg/l de
sodium et moins de 5 mEq/l ou 195 mg/l de potassium.
Le sucrose
permet la reconstitution du glycogène musculaire, alors que le fructose
d'absorption plus lente permet plutôt la reconstitution du glycogène hépatique.
Le fructose est cependant moins bien toléré.
La capacité maximale d'absorption de l'eau
par l'intestin est de 12 ml/kg/h. La meilleure vidange s'effectue lorsque le
liquide est ingéré entre 8 et 13°C.
De faibles quantités de NaCl favorisent la
vidange gastrique. Cependant il est, la plupart du temps, inutile de prendre du
sel car en augmentant le sel on augmente la fuite de potassium. En tout état de
cause, la supplémentation en sel est inutile si la perte d’eau n’atteint pas
2.7 Kg. La supplémentation en glycérol serait meilleure pour
retenir l'eau corporelle.
La vidange gastrique étant limitée, trop
boire revient à surcharger inutilement l'estomac, mais boire insuffisamment
fait courir le risque de la déshydratation.
Il peut se poser des problèmes de
refroidissement trop important quand s'additionnent
- ensoleillement direct (absence de nuages)
- degré hygrométrique bas
- fort vent contraire favorisant le
refroidissement dès l'arrivée
- vitesse de course élevée
Le refroidissement à l'arrivée est très
rapide alors que la production de chaleur chute, et la température corporelle
peut s'effondrer jusqu'au seuil d'hypothermie.
Il est donc préférable de choisir le retour
vent arrière pour freiner le refroidissement et de ne pas arrêter brusquement
l'exercice, mais d'adopter un retour au calme actif.
Les symptômes du coup de chaleur
Les premiers symptômes du coup de chaleur
sont:
Ø
l'arrêt de la sudation
Ø
un pouls rapide et faible
Ø
faible pression sanguine à la station debout
Ø
la " chair de poule " sur les bras et la poitrine
Ø
des frissons
Ø
le vertige
Ø
la nausée
Ø
une température rectale élevée ( la mort survient à 45°C)
Ø
la peau sèche et chaude
Ø
des maux de tête
Ø
des mouvements moins précis
Ø
une faiblesse généralisée
Ø
la confusion
Ø
l'évanouissement
Le coup de chaleur est la plus sérieuse et
la plus complexe des formes de stress thermique. Il résulte essentiellement
d’une défaillance des systèmes thermorégulateurs, provoquée par une très haute
température corporelle. Quand le système thermorégulateur devient défaillant,
le stress s’intensifie jusqu’à causer la mort par défaillance circulatoire et
par atteinte du système nerveux central. Il faut voir très rapidement à
abaisser la température corporelle au moyen de friction à l’alcool,
d’application de sacs de glace ou d'immersion dans l’eau froide.
Le mauvais ajustement circulatoire provient
d’une perte de liquide intracellulaire, surtout du volume plasmatique, réduit à
cause de la sudation excessive. Le sang s’accumule habituellement dans les
vaisseaux périphériques dilatés, ce qui réduit la quantité centrale de sang
nécessaire au maintien du débit cardiaque.
Les soins au sujet victime d'un coup de chaleur
Si un athlète présente des symptômes de coup
de chaleur, il faut immédiatement donner des premiers soins.
Ø
Retirer immédiatement tous les vêtements.
Ø
Abaisser immédiatement la température par n’importe quel moyen disponible:
tuyau d’arrosage, eau glacée, douche froide, application continuelle d'éponges
ou serviettes glacées.
Ø
Faire venir une ambulance et avertir l’hôpital que la personne souffre d’un
coup de chaleur.
Ø
Dans l’ambulance, continuer à abaisser la température à l’aide d’éponges ou de
serviettes trempées dans de l’eau glacée.
§
Il est important d'acclimater progressivement les sportifs à la chaleur, car
l'acclimatation améliore la circulation sanguine et la sudation.
L'acclimatation se produit entre 5 à 8 jours d'entraînement progressif par
grande chaleur. Il est donc nécessaire de s'entraîner quand il fait chaud pour
favoriser l'acclimatation.
§
Le seul fait de se reposer à la chaleur développe très peu de tolérance à la
chaleur. § L'acclimatation doit intervenir avant l'entraînement
proprement dit.
§
L’acclimatation peut être obtenue à l’aide d’un programme d’exercices
progressifs répartis sur 5 à 10 jours, à la chaleur.
§ Un programme d’acclimatation comprend par exemple 20
minutes de travail sous-maximal suivi d’un repos de 20 minutes la première journée.
Tous les jours, la durée de l’exercice, la charge de travail et la quantité de
vêtements sont accrues. La dernière journée, l’exercice peut durer 30 minutes,
la charge de travail peut être maximale et le sujet peut être complètement
vêtu; les périodes de repos durent de 10 à 15 minutes.
§ Un autre programme prévoit un acclimatement à la
chaleur en 4 à 7 jours d'exposition par périodes de 4 heures minimum associé à
un exercice d'intensité progressive, qui sera complet au bout de 12 à 14 jours.
L'élément primordial en est l'accroissement du volume plasmatique.
§ Au cours des périodes de repos, on devrait donner à
boire de l‘eau ou une solution spécifique.
§ L'entraîneur doit surveiller les symptômes d'un
éventuel coup de chaleur et adapter la tenue vestimentaire aux conditions de
l'exercice. Dans une ambiance très chaude, il est conseillé d'espacer des
périodes d'exercice de 20 mn par des périodes de repos de 10 mn.
§
L’acclimatation entraîne une production de sueur plus rapide (37,2°C).
§
Après 10 jours d’acclimatation à la chaleur, la capacité de sudation est
presque doublée. La sueur est plus diluée (moins de sel perdu) et mieux
répartie à la surface de la peau. Chez un individu acclimaté, la perte d’eau
par sudation peut atteindre un maximum de 3 l./h durant un travail intense,
soit en moyenne 12 l./24 h de travail.
§
La différence d'électrolytes dans la sueur peut varier dans la proportion de 3
entre un sujet acclimaté et un autre non-acclimaté (7.29 g/l NA et 2.11 g/l A).
§
La chaleur interne en fin d’exercice peut atteindre 40°. Au cours de l'exercice
maximal, la température rectale peut dépasser 40° et la température cutanée 41°
sans inconfort du sujet.
§
L'efficacité intellectuelle est diminuée au delà de 25°de température ambiante
pour un individu non acclimaté et au delà de 30 à 35° pour un individu
acclimaté.
§
Les aptitudes d'acclimatation ne se détériorent pas avec l'âge.
Le sauna finlandais peut être utilisé dans
la phase d'acclimatation à la chaleur; le déroulement de la séance est composé
de deux phases: la phase de réchauffement et la phase de refroidissement.
Elle se passe dans une pièce dont la
température de l'air est comprise entre 80 et 120°C et le degré d'humidité
entre 5 et 15°. L'idéal reste une température de 80 à 90°C avec un taux
d'humidité relative de 10 à 15%. Cette phase de réchauffement dure de 8 à12
minutes et se renouvelle à peu près trois fois.
Effets
de la phase de réchauffement
u augmentation de la température cutanée et centrale
u sudation
u activation du système sympathique
u élévation de Fc de 50% environ
u augmentation du volume d'éjection systolique
u augmentation de 100% du débit cardiaque
u diminution des résistances périphériques de près de
50%
u pas d'élévation de la tension artérielle chez les
normotendus
u vasodilatation des vaisseaux coronaires
u augmentation de la fréquence respiratoire
u augmentation du volume courant
u accroissement de la dépense énergétique de 20 à 40%
u renforcement des mécanismes de défense
u passage des liquides intracellulaires dans le plasma
pour compenser la perte d'eau (ne pas boire pendant un sauna pour ne pas gêner
ce transfert de liquide et le "décrassage cellulaire")
u perte d'électrolytes infime qu'il est inutile de
compenser par une complémentation alimentaire spécifique
u relâchement de la musculature, effet décontracturant
Elle comporte un passage de 8 à 12 minutes
dans une pièce à air frais, suivi d'une douche ou d'un bain d'eau froide
(dangereux pour les sujets porteurs de problèmes cardiaques ou/et tensionnels)
et de repos.
Effets
de la phase de refroidissement
u chute de la fréquence respiratoire
u diminution de la fréquence cardiaque (plus rapide en
cas de bain avec immersion du visage, qu'en cas de bain sans immersion de la
tête, et plus rapide qu'en cas de douche, car plus de 50% des récepteurs du
froid sont situés dans cette région). Le froid doit être étendu progressivement
de la périphérie vers le cœur pour améliorer le retour veineux.
u saturation du sang en oxygène
u normalisation de la température corporelle
u installation d'un tonus vagotonique avec sensation
de bien-être et de repos
L'ingestion d'une boisson froide après le sauna fait monter PAS à 360 mmHg et PAD à 180 mmHg
u une augmentation de Fc de plus de 60%, du débit
cardiaque (73% chez les sujets sains, 65% chez les hypertendus),
u une diminution légère de la pression artérielle
systolique chez les sujets sains, et importante chez les hypertendus (29 mmHg
en fin de sauna et 50 mmHg vingt minutes après.)
u une diminution de la pression diastolique chez les
hypertendus
u une diminution des résistances périphériques
u maladies inflammatoires
u infections générales
u maladies infectieuses
u infarctus du myocarde aigu
u état de décompensation cardio-vasculaire
u pathologies coronaires
u troubles de la circulation cérébrale
u hypertension > 200 mmHg
u maladies critiques
u hyperthyroïdie
u glaucome
u maladies sexuelles
u tumeurs
u maladies veineuses chroniques
u blessures récentes
Intérêt du sauna pour le sportif
u détente musculaire
u augmentation de la résistance aux maladies
u surcompensation plus rapide des réserves
glycogéniques
u apaisement psychique
u renforcement des capacités de concentration
L'utilisation du sauna pour maigrir et
"faire le poids" conduit à une diminution des capacités sportives.
L'adaptation de l'organisme au froid
Le froid augmente le travail du myocarde et
ses besoins en O2 parce que la vasoconstriction périphérique
destinée à réduire les pertes caloriques augmente les résistances
périphériques, la tension artérielle systémique et diminue le retour veineux.
L'exposition de la figure au froid provoque une bradycardie (réflexe vagal dû à
la stimulation du trijumeau) et une vasoconstriction périphérique. Le froid
favorise la crise d'angor chez le patient coronarien. Le vent possède les mêmes
contre-indications que le froid.
Contre le froid, l'habillement correct est
constitué de plusieurs vêtements (chaque couche de tissu emprisonnant de l'air)
amples pour assurer une circulation de l'air et une ambiance sèche. Toutes les
parties du corps doivent être couvertes y compris la figure et les extrémités
des membres. Pour isoler du froid, les mailles des fibres vestimentaires
emprisonnent de l’air qui se réchauffe. C'est pourquoi plusieurs couches de
vêtements légers ou les vêtements doublés de fourrure, de plumes ou des tissus
synthétiques (formés de plusieurs couches d’air emprisonné) constituent de
meilleurs isolants qu’un seul vêtement d’hiver épais. L'organisme perd environ
30 à 40% de chaleur par la tête. Quand les vêtements se mouillent, ils perdent
90% de leurs propriétés isolantes; le vêtement d’hiver idéal par temps sec est
imperméable à l’air, mais perméable à la vapeur d‘eau de la peau qui transpire.
Un individu moyen ne peut supporter une
immersion supérieure à 20-30 mn dans une eau à 5°C ou à 1 h 30 à 2 h dans une
eau à 15° C. Dans l’eau froide la perte de chaleur par conduction est énorme.
La valeur métabolique d‘un exercice modéré est souvent insuffisante pour
équilibrer les fortes pertes.
La vitesse de conduction nerveuse diminue
avec le froid, mais aussi avec la chaleur à partir d'une certaine température.
Un abaissement de la température cutanée de
quelque degrés peut entraîner un doublement du métabolisme. Le froid augmente
les dépenses de base du sujet maigre, il est inefficace sur un sujet adipeux.
Un surplus de tissu adipeux sous-cutané favorise la performance d’un individu
en ambiance froide. Pour la majorité des individus, c’est entre 26 et 30°C que
l’on se départit le mieux de la chaleur métabolique, sans pour autant geler et
voir la capacité de travail réduite. Au cours d’un exercice intense de courte
durée, une eau plus fraîche pourrait s’avérer avantageuse, surtout pour des
individus plus gras.
Les humains s’adaptent moins bien au froid
prolongé qu’à la chaleur prolongée. A cause de l’importante vasoconstriction
périphérique par temps très froid, la température de la peau et des extrémités peut
s’abaisser jusqu’à un niveau dangereux. Les premiers signes d’un accident par
le froid sont un picotement, un engourdissement des doigts et des orteils ou
une sensation de brûlure au nez et aux oreilles. Le vent est un facteur très
important car par temps venteux, les courants d’air augmentent la perte de
chaleur à mesure que la couche d’air chaude autour de l’organisme est remplacée
par de l’air plus frais.
Le tremblement permet de lutter contre le
froid. Le frisson est la première défense contre le froid. Un individu dormant
dans une ambiance froide sans être réveillé par des frissons va mourir
d'hypothermie.
L'acclimatement au froid se fait par des
expositions brèves et intermittentes à un froid intense. L'exposition répétée
de la face et des mains au froid entraîne une diminution sympathique et un
renforcement de la stimulation vagale.
La récupération après l'effort
La testostérone
accélère tous les mécanismes de resynthèse après effort.
Paiement de la dette d'oxygène
Il faut au moins de 2 à 3 minutes pour que
la consommation d’oxygène (VO2) s’ajuste à l’effort créant ainsi une
dette d’O2. La réserve d’oxygène intramusculaire est reconstituée
entre 10 et 60 secondes.
En cas d'effort modéré et aérobique plus de 50% de la dette d'O2 est
payée en moins de 30 s et en quelques minutes la récupération est complète.
En cas d'effort intense, dépassant la
capacité aérobique maximum, la dette d'O2 est payée de quelques minutes à plus
de 24 h (marathon). La dette d’O2 alactique est rapide et la dette
d'O2 lactique lente
La récupération d'O2
supplémentaire est nécessaire car la restauration des réserves d’O2
attribue 50 ml d'O2 aux muscles respiratoires et au myocarde dont
l’activité reste élevée.
Après effort, la consommation d'oxygène
reste plus élevée qu'à l'état de repos absolu. Le degré et la durée de cette
élévation dépendent de l'intensité de l'effort de l'exercice. La fraction
d'oxygène en sus de celle utilisée habituellement au repos sert à restaurer les
ressources énergétiques.
Remboursement de la composante alactique de la dette d’O2: 3 à 5min
Remboursement de la composante lactique de la dette d’O2: 30 mn à l h
Rétablissement des réserves d’O2: 10-15 s à 1 min
La consommation d'oxygène décroît de 50% en
30 s en association avec une restauration aérobie des réserves d'ATP et CP, et
des réserves d'O2 de l'organisme (myoglobine, hémoglobine).
La consommationd'O2 diminue
encore de 50% pendant une quinzaine de minutes en association avec
l'élimination ou le reconditionnement lactique.
Pour améliorer la capacité du métabolisme
aérobie, la durée de la période de récupération doit être égale ou de 50%
supérieure à celle du travail. Ainsi, au cours d’un exercice de 60 à 90
secondes, l’organisme ne dispose pas assez de temps pour élever sa consommation
d’oxygène de façon à satisfaire les exigences énergétiques de l’exercice.
Le retour à la normale de la consommation
d'O2 peut prendre plus de 60 mn.
L'exercice détermine une augmentation du double de la consommation d'O2 dans l'heure qui suit, et une légère augmentation du métabolisme basal durant au moins 24 heures.
Tous les systèmes physiologiques activés
pour satisfaire les besoins générés par l’exercice augmentent aussi leurs
besoins propres d’oxygène durant la phase de récupération.
La myoglobine ou hémoglobine du muscle est
une réserve musculaire de 500 ml d'O2 qui retarde l'hypoxie et
fournit au muscle les premières mesures d'oxygène nécessaires à la
récupération. La myoglobine se restaure en 1 à 2 minutes.
La récupération des faibles réserves d'ATP
se fait en quelques secondes. Il ne semble pas exister de preuves d'un
mécanisme de surcompensation de l'ATP.
Restauration de la phosphocréatine
Elle a lieu en deux à cinq minutes. 50% est
restauré en 30 secondes, 70% de la restauration s’effectue au cours des 50
premières secondes et le reste dans les 3 à 5 minutes qui suivent. En cas
d’occlusion de la circulation il n’y a aucune récupération des phosphagènes.
Rétablissement des réserves de phosphagènes du
muscle: 2 à 5 mn
La dette alactique est en relation avec la
quantité de phosphagènes utilisés pendant l’effort. Elle peut atteindre 2,5
litres chez l’adulte sédentaire et 6 litres chez le sportif entraîné. D’après
Fox, le remboursement de cette dette s’effectue entre 3 et 5 minutes. Pour des
efforts répétés au cours des périodes de récupération entre les exercices (ou
des périodes de repos d’un exercice intermittent), une partie des réserves de
phosphagènes et d’oxygène intracellulaire est restaurée grâce au système
aérobie. La recharge de la créatine phosphate dépend de l’état interne de la
cellule, en particulier des conditions d’acidose, qui en augmentent
sensiblement la durée (70% seulement de la quantité initiale en 2 à 3 minutes).
En pratique, il convient donc de prendre
garde
ð aux échauffements hâtifs ou mal dosés qui acidifient
prématurément la musculature. L’acidose provoque un allongement de la durée de
resynthèse de la créatine phosphate, et une inhibition de la dégradation des
phosphagènes. Leur puissance d‘utilisation en est réduite
ð aux répétitions d’efforts susceptibles de stimuler
fortement la glycolyse anaérobie
La période de récupération peut être passive
(repos) ou active (travail léger). Une période de récupération trois fois plus
longue que celle du travail convient bien à l'entraînement en puissance
anaérobie alactique. Ainsi, un sprinter qui court 10 secondes devrait se
reposer 30 secondes. Pour améliorer la capacité de la glycogénolyse anaérobie,
il faut allouer deux fois plus de temps de récupération que de travail.
Le respect de l’intensité du travail de
vitesse maximale de course réclame des durées de récupération suffisante afin
de ne pas activer trop fortement les processus lactiques. Quatre séries de
quatre sprints de 40 mètres sont effectuées avec des récupérations d’une minute
entre les courses, de quatre minutes entre les séries. chez des footballeurs de
17 ans. Les résultats montrent que seize efforts très brefs de 5,4 s en 24
minutes stimulent la glycolyse anaérobie (lactatémie sanguine à 8 mmol/l).
On connaît moins les capacités de
surcompensation de la phosphocréatine, cependant il apparaîtrait que le pic de
surcompensation de la phosphocréatine survient beaucoup plus tôt que celui du
glycogène, pour revenir à l'état initial au moment où le glycogène est à son
maximum. Cette notion est importante, car elle tendrait à prouver la valeur
d'un entraînement dans les 24 heures précédant une compétition pour les sports
de la filière anaérobie alactique (ATP + CP). On peut logiquement penser que le
dernier entraînement devrait être pratiqué quelques minutes (10 à 15) dans la
filière ATP + CP, en mixant des exercices très courts et très violents (@ 6 s) et des
temps de repos de moins de 30 secondes pour user les réserves ATP et CP en
préservant les réserves glycogéniques. Il serait néanmoins intéressant
d'affiner les rapports de la surcompensation en CP avec l'intensité et la durée
du dernier entraînement, les délais d'obtention du pic de surcompensation en CP
et le degré d'usure et de récupération du glycogène après cet exercice.
Une étude présentée par Jakovlev semble
démontrer un pic de surcompensation intéressant de CP 6 h après un exercice de
15 minutes. A ce moment-là le glycogène est également en état de
surcompensation, même si il n'a pas atteint son pic (12 h). Il en est de même
pour le métabolisme protéique qui atteindra son pic de surcompensation dans ces
conditions après 12 h. Les pics de surcompensation précèdent la phase de retour
aux valeurs initiales.
Resynthèse du glycogène musculaire :
ð après un exercice continu 10 h à 48 h
ð après un exercice par intervalles 5 h à 24 h
Resynthèse des réserves de glycogène hépatique: inconnu à 12-24 h
La restauration de glycogène débute dès la première heure qui suit
l'exercice, elle est reconstituée à 50% après 10 à 12 heures, elle demande 48 à
72 heures pour être complète en fonction des individus.
Le degré d'usure des réserves de glycogène
conditionne la durée de restauration complète.
Exercices
épuisants de longue durée
L'exercice continu dont le substrat
principal est l'acide gras libre, use beaucoup plus les réserves de glycogène
que l'exercice intermittent dont le substrat énergétique principal est
l'hydrate de carbone donc le glycogène. Il est nécessaire de se rappeler que
l'exercice aérobique lipolytique ne peut avoir lieu qu'en présence de
glycogène, et qu'à un certain seuil la déplétion de glycogène interrompt cet
exercice. Alors même que l'exercice intermittent est réputé parce qu'il permet
des intensités plus élevées avec une moindre accumulation d'acide lactique, on
peut imaginer que l'exercice fractionné génère une fatigue ou une accumulation
de protons telles que l'organisme doit cesser l'exercice avant d'avoir usé
suffisamment de glycogène, comparativement à l'exercice aérobique.
Il faut noter que l'exercice sous
électrostimulation est celui qui permet de puiser le plus dans les réserves
glycogéniques d'un muscle. Ceci est
du au fait que le travail est pratiqué en mode réflexe par la stimulation des
fibres Ia et Ib, et que le circuit de protection de Renshaw est inhibé dans ces
conditions. En fait, là ou la sensation de fatigue intervient pour arrêter
l'exercice volontaire, l'électrostimulation impose la poursuite du travail.
L'organisme garde toujours une fraction de glycogène intacte pour permettre un
effort vital si nécessaire (loi de la conservation) dans laquelle va puiser
l'entraînement sous électrostimulation. Ce mécanisme est renforcé par
l'antalgie qu'exerce l'électrostimulation et qui masque la fatigue et les
douleurs.
Il existe la possibilité d'obtenir une
augmentation des réserves de glycogène par surcompensation. Il faut pratiquer
un régime légèrement hypoglucidique avant de pratiquer une série d'exercices
aérobiques, ou mieux sous électrostimulation, destinée à épuiser les réserves
de glycogène. Cette série d'exercices s'étale sur trois jours. Le patient est
ensuite mis au repos, ou sous activité légère pendant deux à trois jours avec
une alimentation hyperglucidique. On arrive ainsi à augmenter les réserves de
glycogène de 250%. La surcompensation la plus spectaculaire se produit au
niveau des muscles utilisés dans l'exercice.
Les études ont montré qu’une très petite
quantité de glycogène musculaire est resynthétisée au cours de la période
initiale (1 à 2 heures) qui suit un exercice d’endurance de 2 heures; la
resynthèse complète du glycogène musculaire après un exercice d’endurance
nécessite un régime riche en glucides pendant les 2 jours de récupération. En
effet, seule une petite partie du glycogène est resynthétisée, même après une
période de 5 jours, si l’apport de glucides est nul. De plus, ces études ont
montré que le réapprovisionnement du glycogène musculaire en présence de
glucides s'effectue plus rapidement au cours des premières heures de
récupération (après seulement 10 heures, la resynthèse est complétée à 60 pour
cent). Le principal responsable du rétablissement de la concentration initiale
de glycogène est le sucre alimentaire et non la resynthèse de l’acide lactique.
Un apport de glucose est donc indispensable à la resynthèse du glycogène. Une
alimentation hyperglucidique est également indispensable dans les efforts
d'endurance portant sur plusieurs jours consécutifs. En effet, les réserves de
glycogène sont progressivement épuisées au cours de 3 jours de course à raison
de 16 km/jour et ce, malgré une ingestion normale de glucides au cours de cette
période.
Exercices épuisants de courte durée
Il est prouvé:
u qu’une quantité importante de glycogène peut être
resynthétisée pendant la partie initiale de la récupération, même si aucune
nourriture n’est absorbée;
u que la resynthèse complète du glycogène musculaire
ne nécessite pas un apport supplémentaire de glucides et qu’elle est terminée
après 24 heures de récupération;
u que la resynthèse du glycogène musculaire est plus
rapide au cours des premières heures de récupération (elle est rétablie à 39%
en 2 heures et à 53% en 5 heures).
Explications
La quantité totale de glycogène utilisée au
cours de l’exercice continu est environ deux fois plus grande que celle
utilisée au cours de l’exercice intermittent. Un autre facteur important a
trait à la disponibilité des précurseurs du glycogène. La synthèse du glycogène
implique la présence de quantités adéquates de ces précurseurs dont les
principaux sont l’acide lactique, l’acide pyruvique et le glucose. Ceux-ci
doivent être présents au niveau du foie et du muscle où la majeure partie de Ia
resynthèse s'effectue. Après un exercice continu prolongé, la plupart de ces
précurseurs sont présents en quantité réduite. Par contre, l’exercice
intermittent n’entraîne pas de diminution de ces substrats et la resynthèse du
glycogène peut débuter plus tôt dans ce cas. De plus, ce phénomène explique
pourquoi l’absorption de glucides est essentielle après un exercice continu,
mais non à la suite d’un exercice intermittent et pourquoi une plus grande
ingestion de glucides n’accélère pas la resynthèse du glycogène après un
exercice par intervalles. De plus il a été montré que la resynthèse du
glycogène dans les fibres FT est plus rapide que dans les fibres ST.
Elimination de l'acide lactique
 Après un exercice intense le taux de lactate sanguin est normalisé
environ une heure après l'exercice. La normalisation est améliorée par un
exercice aérobique à 40% de Fc Max. Après un exercice maximal, un individu au
repos complet élimine 50% de l'acide lactique en 15 mn, environ 88% en 75 min
de récupération.
Après un exercice intense le taux de lactate sanguin est normalisé
environ une heure après l'exercice. La normalisation est améliorée par un
exercice aérobique à 40% de Fc Max. Après un exercice maximal, un individu au
repos complet élimine 50% de l'acide lactique en 15 mn, environ 88% en 75 min
de récupération.
L‘intensité de l’exercice de récupération
qui entraîne l’élimination la plus rapide de l’acide lactique correspond à
environ 30 à 40% du V02 Max.
Chez des sujets entraînés, on a montré que l’élimination de l’acide lactique
est plus rapide lorsque la récupération s’effectue à une intensité d’environ 50
à 60% du VO2 Max.
Si l’intensité est supérieure à 60% du VO2
Max, l’élimination de l’acide lactique est plus lente qu’au cours de la
récupération inactive car il y a de nouveau production d’acide lactique.
L'acide lactique est excrété pour petite
partie dans l'urine et la sueur. L'acide lactique est transformé en H2O
+ CO2, en pyruvate, en glycogène musculaire (50 à 75%) ou en acide
pyruvique, précurseur du glucose hépatique oxydé dans le cycle de Krebs.

récupération active : 30 min à 1 h
récupération passive : 1 h à 2 h
Il est double:

¨ la moitié reste dans la cellule
¨ la moitié sort de la cellule avec plusieurs
destinées:
¨ passage dans le liquide extracellulaire où il est
tamponné
¨ par la voie sanguine, il atteint le myocarde qui en
est un grand consommateur
¨ par la voie sanguine, il atteint le foie et le rein
où il est transformé en glucose
¨ par la voie sanguine, il atteint le rein qui
l’élimine mais la quantité en est très faible
¨ il peut revenir dans la cellule musculaire au repos
en situation aérobique.
L’acide lactique métabolisé est à 65% oxydé
en CO2 et H20, à 25 % resynthétisé en glucose , et à l0%
resynthétisé en protéines.
La demi-vie de l'acide lactique est de 25 mn.
L'acide lactique post-exercice est donc éliminé en une heure,
préférentiellement lors d'une récupération active en aérobie.
Boire légèrement avant la course, permet de
déclencher la transpiration plus tôt et donc de mieux refroidir; cependant
c'est un risque de gaspillage des réserves si l'effort doit être accompagné
d'une grande déshydratation.
L'ingestion de sucres rapides (index
glycémique élevé) dans l'heure qui précède la compétition entraîne une
hyperinsulinémie. Or l'insuline inhibe la lipolyse et s'opposera à une
utilisation optimale des acides gras pour épargner le glycogène. De plus
l'hyperglycémie provoquera une hypoglycémie réactionnelle, et donc un coup de
fatigue.
 L'ingestion de caféine à haute dose (200 à 300 mg = 3 tasses), dans
l'heure qui précède et si elle est supportée permet de privilégier la lipolyse
et l'utilisation des acides gras. L'héparine tient le même rôle, mais est
dangereuse par son effet anticoagulant. La caféine est un produit dopant (12
mg/l = 10 tasses).
L'ingestion de caféine à haute dose (200 à 300 mg = 3 tasses), dans
l'heure qui précède et si elle est supportée permet de privilégier la lipolyse
et l'utilisation des acides gras. L'héparine tient le même rôle, mais est
dangereuse par son effet anticoagulant. La caféine est un produit dopant (12
mg/l = 10 tasses).
La caféine possède des inconvénients par son
effet excitant, et stimulant du métabolisme intestinal.
 L'alcool possède le pouvoir de désinhiber, mais également celui de
déshydrater, en freinant la production de l'hormone antidiurétique par les
reins.
L'alcool possède le pouvoir de désinhiber, mais également celui de
déshydrater, en freinant la production de l'hormone antidiurétique par les
reins.
Le dernier repas avant l'effort doit être
pris trois à quatre heures avant le début de l'exercice, de manière à ne pas
être gêné par la digestion. Le dernier repas doit être à prédominance
glucidique, dont une partie en sucres lents et en fructose. Il vaut mieux
éviter les protéines et les graisses qui se digèrent plus lentement et
n'apportent pas d'énergie exploitable pendant l'exercice. Il vaut mieux
préférer un repas à base de pommes de terre cuites à l'eau dans leur peau ou de
pâtes. Les pommes de terre semblent mettre le sportif à l'abri d'une
défaillance dans la première demi-heure d'effort mieux que les pâtes. Le repas
de la veille au soir reste conseillé à base de pâtes pour mieux renforcer les
réserves glycogéniques.
Les pâtes en repas pré-compétition, à la
différence de la pomme de terre ne mettent pas à l'abri d'une hypoglycémie lors
des 30 premières minutes d'effort. La préparation et la cuisson de la pdt
jouent sur son index glycémique. Plus la pdt est coupée ou broyée, et plus elle
est chauffée, plus son index glycémique s'élève. Sa peau est source de
nombreuses vitamines et agents. La pdt cuite à l'eau 20 mn avec sa peau tient
un index glycémique proche de celui des pâtes. En conséquence si le repas de la
veille doit être à base de pâtes, celui d'avant compétition est préférable à
base de pdt.
Il faut manger avant l'exercice pour
surcharger le foie en réserve de glycogène. Il est déconseillé d'ingérer des
sucres rapides dans les deux heures qui précèdent l'exercice pour éviter une
hypoglycémie en début de travail. Le sujet peut commencer à ingérer du glucose
15 minutes avant le début de l'exercice, car ce glucose sera utilisé par les
muscles.

L'alimentation pendant la compétition

Pour les mêmes raisons et parce que le
glucose s'oppose à la pénétration de l'eau dans le plasma, la boisson absorbée
pendant la compétition devra avoir les caractéristiques suivantes:
L'eau ingérée doit être 25 cl (12 ml/kg/h) d'une solution glucosée à 25 g/l (½ sucrose, ½ fructose) d'une température de 10 à 15°C toutes les quinze minutes et de moins de 10 mEq/l ou 230 mg/l de sodium et moins de 5 mEq/l ou 195 mg/l de potassium.
Le sucrose permet la reconstitution du glycogène musculaire, alors que le fructose d'absorption plus lente permet plutôt la reconstitution du glycogène hépatique. Le fructose est cependant moins bien toléré.
L'ingestion de sucre juste avant la fin de
l'effort, permet de privilégier la réserve de glucose, plus explosive, au
détriment des acides gras pour accélérer ou sprinter.
Les supplémentations, notamment en sel
semblent inutiles, car elles tendent à concentrer les solutions du fait de la
plus grande perte d'eau.
Il doit avoir lieu 1 h 1/2 après la fin de
l'exercice et être essentiellement composé d'aliments à index glycémique élevés
(purée, pâtes …) pour reconstituer les réserves de glycogène. On préférera la
purée qui possède un index glycémique supérieur, favorisant une meilleure
reconstitution des réserves de glycogène aux pâtes ou aux pommes de terre
cuites à l'eau.
La meilleure reconstitution du glycogène se
produit avec un repas post-exercice dans les deux heures qui suivent et un
deuxième repas dans les deux à quatre heures suivant ce repas. Une charge
glucidique égale à 70% de la dépense calorique de l'exercice constitué
d'aliments à index glycémique élevé (sucres rapides) pour une proportion entre
1/3 à1/2 afin d'éviter les "chocs insuliniques" qui ont tendance à
utiliser ces sucres pour constituer des dépôts adipeux.
La récupération post-exercice nécessite 50 g
d'hydrate carbone par 2 h.
Causes de la douleur musculaire et de la raideur
ð micro-déchirures du tissu musculaire
ð modification de la pression osmotique causant une
rétention d’eau dans les tissus avoisinants
ð spasmes musculaires
ð surétirement et déchirure de portions du tissu
conjonctif intramusculaire
ð libération d’histamine et de polypeptides vasodilatateurs
provoquant un œdème douloureux par la micro-compression des terminaisons
nerveuses
sont essentiellement dues à l'accumulation
de métabolites, à un œdème interstitiel et à des micro-lésions des structures
musculaires au niveau des ponts actine-myosine. Il faut noter que les
contractions pliométriques causent plus de lésions ultrastructurales que les
contractions miométriques.
Ces problèmes sont restaurés en 1 à 5 jours
en fonction de l'importance des courbatures. Contradictoirement les courbatures
dues à un petit exercice sur des muscles non entraînés à cet exercice procurent
plus de douleurs et mettent plus de temps à se résorber que les courbatures
dues à un exercice d'intensité et de durée importantes sur des muscles entraînés
à cet exercice. Il suffit de faire répéter le test de Ruffier jusqu'à
épuisement (quelques fois suffisent) à un sportif non entraîné à pratiquer les
flexions des genoux (squats) pour s'en apercevoir.
Les courbatures d'effort sensibles au bout
de quelques heures après l'exercice, deviennent plus importantes le deuxième
jour. Elle disparaissent 4 à 5 jours après l'exercice. Les courbatures touchent
essentiellement les fibres rapides. Le niveau d'apparition des courbatures
augmente avec l'entraînement.
Ne pas masser des courbatures (source
d'irritation mécanique), utiliser chaleur et électrostimulation pour augmenter
la vascularisation et donc la cicatrisation.
Elle possède
différentes étiologies
notamment celle ci:
L'acidose entraîne une entrée cellulaire de
H+, et une sortie de K+. Le catabolisme glucidique et protidique entraîne une
sortie cellulaire de K+ et une augmentation plasmatique de K+.
La régulation se fait par l'excrétion dans
les reins de K+ grâce à l'aldostérone contre réabsorption de Na+ ou H+. La
diminution de Na ou/et l'augmentation de K+ entraînent une sécrétion
d’aldostérone pour diminuer sortie Na et augmenter sortie K+.
On peut éventuellement prévenir la crampe
par une supplémentation de sels de potassium à l’exercice. La supplémentation
en NaCl ne se conçoit que pour se prémunir de pertes importantes d'eau.
La crampe se solutionne par un étirement
(réflexe myotatique inversé de Sherrington).
Un œdème d'effort douloureux peut aussi
survenir sans aucune gravité. En effet si après des exercices anaérobies
intenses, le sujet n'élimine pas rapidement suffisamment d'acide lactique grâce
à une récupération active, l'osmolarité n'est pas rétablie et les cellules
musculaires se gonflent d'eau entraînant une raideur.
est un étirement passif du muscle au-delà de
sa limite physiologique. Le muscle devient douloureux à l’effort et l’examen
montre une douleur à la palpation; après 3 à 10 jours, il y a sans traitement
guérison sans séquelle.
Est une rupture de la fibre musculaire
d'intensité plus ou moins importante en fonction du nombre de fibres touchées.
Elle peut survenir sur l'agoniste au cours de l'effort ou sur l'antagoniste
quand il manque de force ou/et de souplesse et ne peut contrôler et résister à
la contraction de l'agoniste.
L'hypoglycémie se caractérise par :
£ Une sensation anormale de faim impérieuse avec
fringale et état nauséeux
£ Une sensation d’épuisement
£ Des signes objectifs:
£ sueurs,
£ vertiges,
£ confusion mentale,
£ lipothymie,
£ défaillance physique,
£ tachycardie
£ si on dose la glycémie au moment de l’accident on
trouve un taux autour de 0,50 g/l.
Prévention
du syndrome hypoglycémique:
£ l’alimentation avant l’exercice musculaire
£ l’alimentation pendant l’exercice musculaire soit sous forme de glucides rapides (200 ml d’une solution de glucose à 5 % toutes les 15 minutes dans une épreuve à 70 % de la puissance maximum), soit sous forme de glucides lents.
Le traitement curatif du syndrome
hypoglycémique:
£ absorption de glucides rapides, surtout sous forme liquide plus facile à prendre si le sujet est au bord du malaise.
Dans les cas graves:
£ injection intraveineuse d’une solution glucosée à 5%
£ injection sous-cutanée ou intramusculaire de glucagon qui est l’hormone mobilisatrice du glycogène hépatique.
Se caractérise par des sueurs, des nausées,
une bradycardie. Le patient s'effondre. Plus la fréquence cardiaque est montée
haut durant l'exercice et plus la descente du rythme cardiaque a été rapide en
fin d'exercice, plus augmente le risque de malaise vagal. Le retour au calme
actif est un moyen de prévention du malaise vagal.
Mettre le patient dans une position comme
sur la photo ci-dessous ou jambes surélevées et attendre qu'il récupère. Devant
un malaise vagal inexpliqué (atmosphère chaude, choc émotionnel, grossesse,
manœuvre de Valsalva…) pratiquer un ECG.
Troubles
cardiaques pendant l'exercice
Insuffisance cardiaque aiguë
des cœurs sains
trois hypothèses
sont avancées:
è épuisement des réserves glycogéniques de tous les
organes (Frey)
è insuffisance cortico-surrénalienne après une
affection saisonnière (Plas)
è forme d'œdème aigu du poumon.
Troubles du fonctionnement du
tissu nodal
Le professeur Plas estime que ces troubles
atteignent 12 % des sportifs et propose la classification suivante:
è Troubles de conduction observés au repos
è bloc sino-auriculaire
è bloc auriculo-ventriculaire du 1er degré
è dissociation isorythmique
Ces troubles sont bénins sans trouble
fonctionnel et sans risque vital; d’une manière paradoxale on peut dire que les
cœurs des athlètes atteints ne sont normaux que pendant l’action.
è Troubles de l’excitabilité observés pendant l’effort
pouvant provoquer une lipothymie voire un collapsus passager.
è Les tachycardies paroxystiques soulèvent un problème
difficile d’un rythme ectopique: bénin à l’étage auriculaire, il peut être
grave à l’étage ventriculaire
è Les extra-systoles ont un pronostic très différent
suivant leurs caractéristiques.
è Le syndrome de Wolf-Parkinson-White est dû au
passage de l’influx dans une voie anormale et provoque des crises de
tachycardie paroxystique à l’effort.
è Le syndrome du cœur forcé par le surentraînement
provoque une tachycardie avec inaptitude à l’effort comme s’il y avait
disparition du frein vagal alors que le cœur fonctionne normalement au repos.
Les problèmes de !a mort subite du sportif après
autopsie blanche (négative)
è inhibition par excitation du nerf pneumo-gastrique
qui est cardio-modérateur
è brusque excitation externe par un traumatisme local
(coup brutal sur le xiphoïde, la région abdominale, le cou)
è traumatisme global (joueur de rugby pris en "
sandwich " entre 2 joueurs)
è excitation interne par une hémorragie minime (qui
peut passer inaperçue à l’autopsie) dans un territoire riche en fibres
nerveuses
è insuffisance cortico-surrénalienne pouvant
apparaître au décours d’une infection banale traitée dans les semaines qui
précèdent.
Le pseudo-collapsus cardio-périphérique du sportif
Au cours de l'effort, le coureur ralentit
puis s'effondre ou bien à l’arrivée, le coureur s’abat sur le sol. Dans les
deux cas, la respiration est rapide, superficielle, haletante, le pouls très
rapide quasi imprenable, la tension artérielle effondrée puis progressivement
les symptômes s’améliorent. L’hypothèse explicative serait un dérèglement du
contrôle tensionnel issu du corpuscule carotidien aggravé par l’action de
l’hypercapnie sur les centres bulbaire cardio-respiratoires. L‘évolution
spontanée est favorable et ne nécessite aucune thérapeutique.
Physiopathologie
de la mort subite du sportif
La mort subite du sportif résulte la plupart
du temps à l'âge jeune, de rétrécissement aortique, de myocardiopathie
hypertrophique (+ 50%), d'arythmies ventriculaires primaires dont la dysplasie
arythmogène droite; après 40 ans la maladie coronaire athéroscléreuse est
prédominante dans la mort subite du sportif; il s'agit essentiellement de
thrombose sur plaque d'athérosclérose coronaire fissurée, la douleur étant
masquée par les endorphines produites lors de l'effort. Les temps fort du
risque de mort subite du sportif, sont le début de l'exercice, l'arrêt de
l'exercice, alors que la production d'adrénaline va se poursuivre pendant trois
minutes, ou l'heure qui suit l'effort qui a fragilisé la plaque d'athérome.
Les
manœuvres vagales à utiliser en cas de tachycardie (flutter, WPW…)
- compression unilatérale d'une carotide
interne par une compression antéro-postérieure de façon à l'appuyer sur le plan
musculaire paravertébral après s'être assuré à l'auscultation qu'il n'existe
pas de souffle carotidien; si possible surveiller le rythme à l'ECG; changer
éventuellement de carotide, mais ne jamais masser les deux carotides
simultanément;
- compression des globes oculaires qui
doivent être comprimés latéralement en plaçant les pouces dans l'angle interne
de chaque œil et en refoulant les globes oculaires vers la paroi externe de
l'orbite; s'assurer au préalable que le patient ne souffre pas de glaucome et
ne porte pas de lentilles de contact; la compression doit être progressive et
douloureuse pour être efficace;
- manœuvre de Valsalva
- blocage respiratoire
L'efficacité se traduit par une diminution
de 5 bats/min.
Attitude à adopter en cas de malaise cardiaque du patient
Répond-il ? Respire-t-il ? Sentez-vous
son pouls ? Action
oui
oui oui A
non
oui oui B
non
non oui C
non
non non D
Action
A :
F Ne changez pas sa position, protégez-le
F Appelez les secours
F Surveillez-le
Action
B
F Installez-le en position latérale de sécurité, avec
la tête basculée en arrière
F Faîtes attention à un éventuel dégât de la colonne
pour un traumatisé
F Appelez les secours
Action
C
F Libérez les voies aériennes
F Vérifiez l'absence de corps étrangers dans la bouche
F Basculez prudemment la tête en arrière en soulevant
le menton pour ouvrir les voies respiratoires
F Faîtes 2 insufflations en bouche à bouche ou bouche
à nez, en soufflant lentement deux minutes chaque fois
F Appelez les secours
F Continuez les insufflations au rythme de 10 par
minute
action
D
 F Appelez ou faîtes appeler immédiatement les secours
F Appelez ou faîtes appeler immédiatement les secours
F Débutez immédiatement la réanimation
cardio-pulmonaire
F Libérez les voies aériennes et faîtes 10
insufflations par minute
F Si vous avez appris, associez un massage cardiaque
externe en faisant 2 insufflations pour 15 compressions en 15 secondes
F Si vous êtes à deux, l'un fait 1 insufflation
pendant que l'autre fait 5 compressions (toujours au rythme de 60 compressions
par seconde)
F Ne vous arrêtez pas jusqu'à l'arrivée des secours
Précautions
à prendre durant la réanimation cardio-pulmonaire
- Desserrer
les vêtements et la ceinture de manière à libérer les voies aériennes
- Basculer la tête du patient en arrière
- Bien pincer le nez
- Souffler de façon progressive
- Garder le rythme de compression du thorax
à 60 bats/min; souvent le rythme est trop lent
- Ne pas masser trop fort (risque de
fractures des côtes)
- Ne pas masser trop rapidement pour ne pas
se fatiguer
- Bien placer les mains l'une sur l'autre
sur le cœur
- Ne pas souffler l'air dans l'estomac, ce
qui risque d'entraîner une régurgitation du patient entraînant dégoût et
perturbation du sauveteur, les matières régurgitées risquant en outre de
s'introduire dans les poumons
Troubles du système épuratoire dus à
l'exercice
Le mécanisme serait le suivant: chocs
répétés des globules rouges sur une région de surface réduite et hyperhémiée
(congestionnée) telle que plante du pied ou paume de la main induisant le
passage de l‘hémoglobine dans le plasma. Au dessus de 100 mg d’hémoglobine dans
100 ml de plasma, le passage se fait dans l’urine: c’est-à-dire que
l’hémoglobinurie témoigne d’une hémolyse.
Les sports en cause sont: la marche rapide
de longue durée sur un sol dur, la course à pied depuis le demi-fond jusqu’au
marathon, la pelote basque (c’est à dire à main nue) exceptionnellement le
basket-ball, le tennis. Le seul signe est l’urine colorée en rouge et qui
disparaît spontanément en quelques heures; en raison du caractère bénin il faut
s’abstenir de tout traitement.
Myoglobinurie d’effort et rhabdomyolyse
Au cours de l’entraînement ou de l’épreuve
mettant en jeu des groupes musculaires importants, on peut avoir une souffrance
aiguë du muscle avec myolyse libérant de la myoglobine passant dans le plasma ;
au-dessus de 20 mg de myoglobine dans 100 ml de plasma il y a passage dans
l’urine. Les sports en cause sont la course à pied, le football américain, la
lutte, le ski, le patin à glace. Le mécanisme physio-pathologique est inconnu
alors que le tableau clinique est grave: syndrome musculaire avec muscles
gonflés, douloureux et paralysés, syndrome général de malaise important voire
état de choc.
L'évolution est souvent favorable spontanément
mais on a signalé des complications rénales allant jusqu’à l’anurie. Le
traitement banal consiste dans le repos absolu avec l’absorption d’une grande
quantité de liquide. Dans les formes sévères on doit recourir à la réanimation
pouvant comprendre l’hémodialyse. La prévention repose sur la progressivité de
l’entraînement et l’absorption d’une quantité d’eau en fonction de l’effort
musculaire et des conditions atmosphériques.
Troubles du système tampon dus à
l'exercice
La baisse des bicarbonates résulte soit
d’une fuite excessive des bases, soit d’une production excessive d’acides
(jeûne prolongé, gros travail musculaire).
L’élévation des bicarbonates peut résulter
de 3 causes:
è l’excès d’ingestion d’aliments ou de médicaments
basiques: régime végétalien
è prise de bicarbonate de sodium contre les douleurs
gastriques
è les vomissements qui provoquent une perte d’acide
chlorhydrique
è la déplétion potassique par diarrhée: les ions H+ quittent
le liquide extra-cellulaire pour remplacer l’ion potassium du liquide
intracellulaire.
L‘élévation de l’acide carbonique résulte
soit de l’hypoventilation alvéolaire soit de l’asphyxie.
L’acidose d’effort entraîne:
è une diminution de la production d’A.T.P. par
inhibition des réactions de dégradation du glucose en acide pyruvique
è une réduction de la contractilité musculaire, par la
baisse de l’interaction actine/myosine.
La baisse de l’acide carbonique est causée
soit par l’hyperventilation alvéolaire soit par le mal des montagnes.
Fractures spontanées (ou de fatigue)
du sportif
Ce sont des fractures non traumatiques qui
se rencontrent surtout dans trois localisations :
à La fracture de fatigue des métatarsiens atteint le 2ème
et parfois le 3ème métatarsien. La fracture se consolide sans
séquelle par une immobilisation de 4 semaines.
à La fracture de fatigue du péroné se rencontre
surtout chez les coureurs et aussi les parachutistes. Il y a une douleur
progressive pendant le mouvement et qui disparaît au repos; à l’examen, on a un
gonflement local et une gêne à la mobilisation. Les signes radiologiques
n’apparaissent qu’une ou deux semaines après le début clinique avec un trait de
fracture situé entre 4 à 7 cm de la pointe de la malléole péronnière. La
fracture se consolide sans séquelle par une immobilisation de 6 semaines.
à La fracture de fatigue du tibia est rare et se
rencontre chez les danseurs où elle se manifeste par une douleur de la partie
moyenne de la crête tibiale avec gonflement.